

Structure of a sedoheptulose 7-phosphate cyclase: ValA from Streptomyces hygroscopicus
- PMID: 24832673
- PMCID: PMC4095911
- DOI: 10.1021/bi5003508
Structure of a sedoheptulose 7-phosphate cyclase: ValA from Streptomyces hygroscopicus
Erratum in
- Biochemistry. 2014 Jul 8;53(26):4316
Abstract
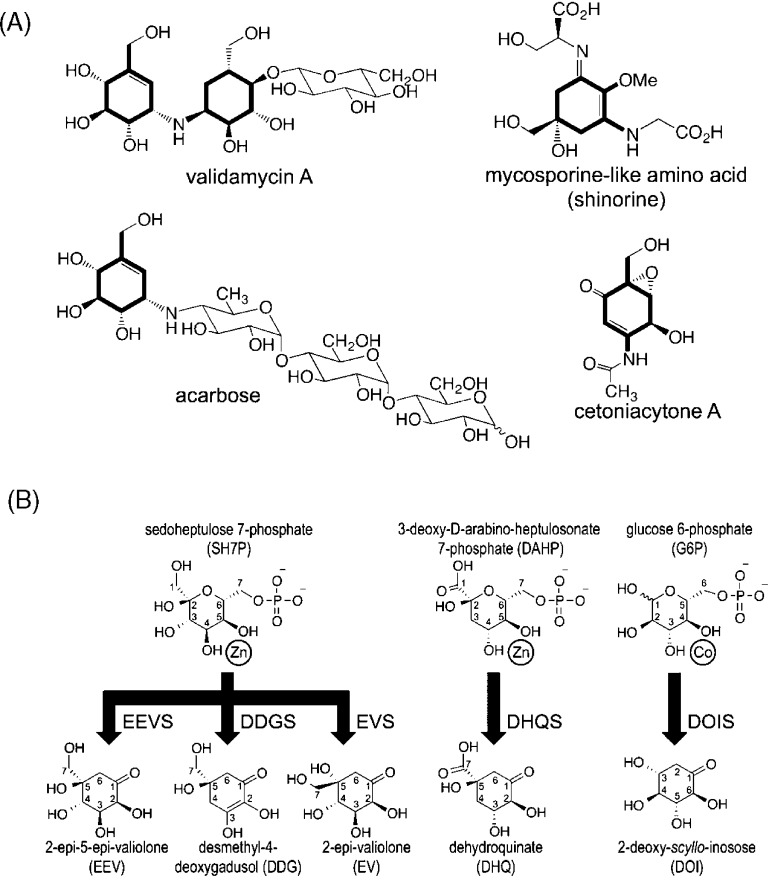
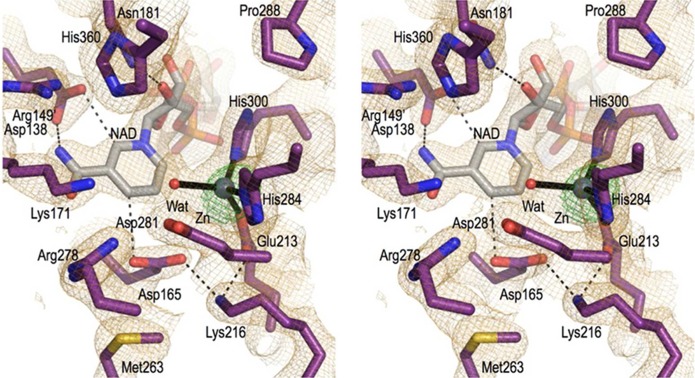
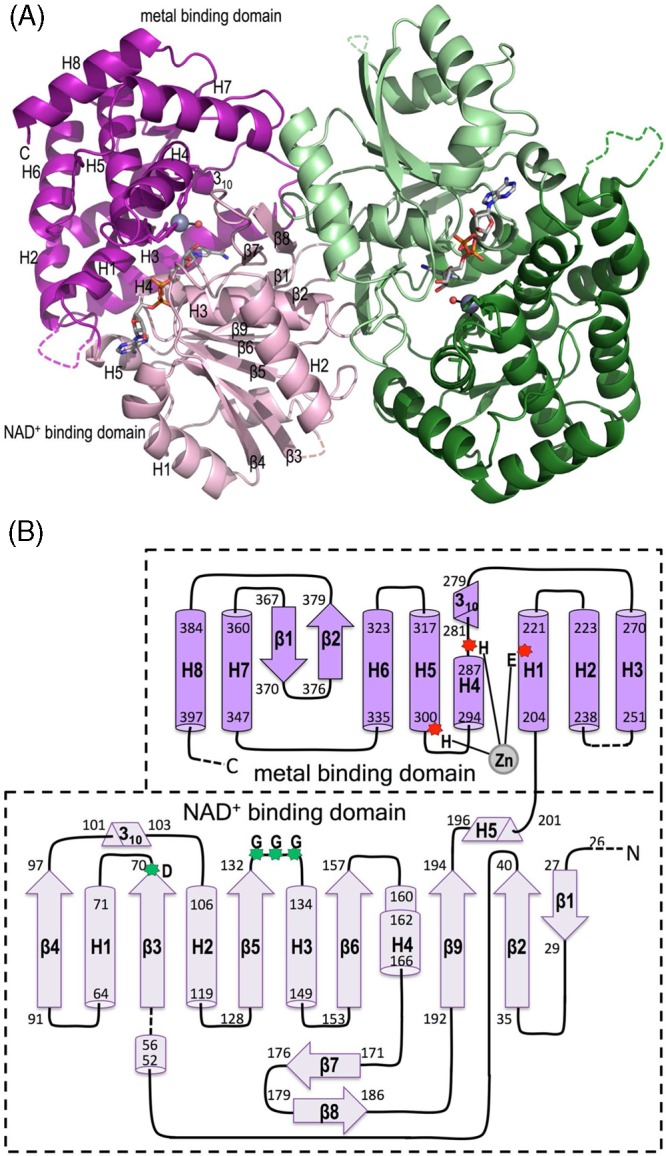
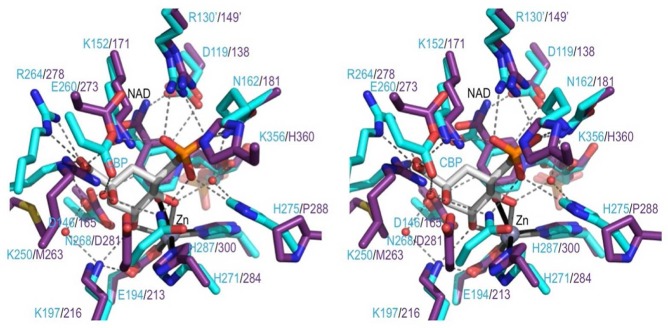
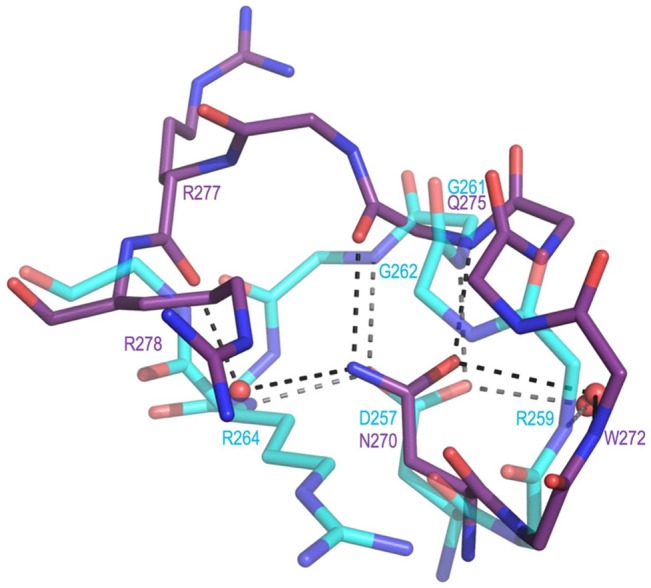
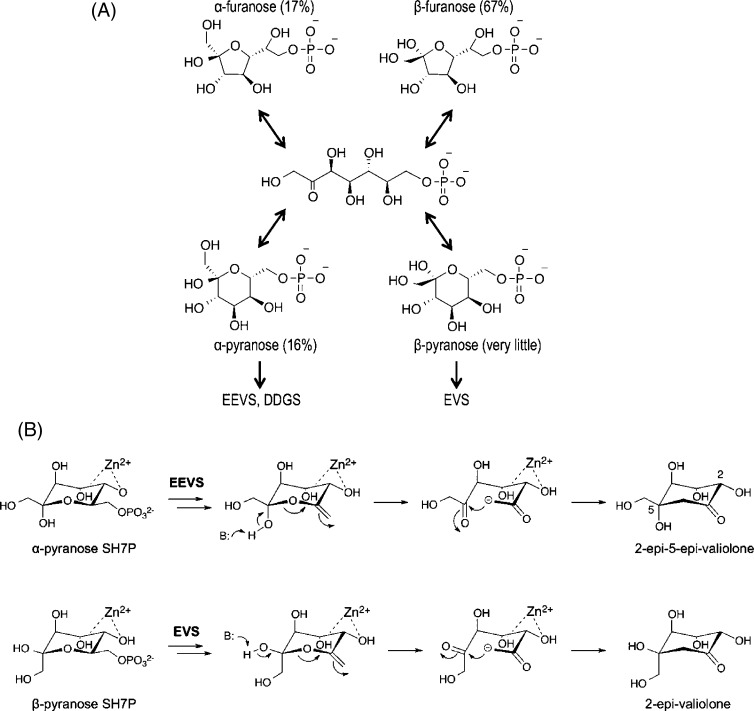
Sedoheptulose 7-phosphate cyclases (SH7PCs) encompass three enzymes involved in producing the core cyclitol structures of pseudoglycosides and similar bioactive natural products. One such enzyme is ValA from Streptomyces hygroscopicus subsp. jinggangensis 5008, which makes 2-epi-5-epi-valiolone as part of the biosynthesis of the agricultural antifungal agent validamycin A. We present, as the first SH7PC structure, the 2.1 Å resolution crystal structure of ValA in complex with NAD+ and Zn2+ cofactors. ValA has a fold and active site organization resembling those of the sugar phosphate cyclase dehydroquinate synthase (DHQS) and contains two notable, previously unrecognized interactions between NAD+ and Asp side chains conserved in all sugar phosphate cyclases that may influence catalysis. Because the domains of ValA adopt a nearly closed conformation even though no sugar substrate is present, comparisons with a ligand-bound DHQS provide a model for aspects of substrate binding. One striking active site difference is a loop that adopts a distinct conformation as a result of an Asp→Asn change with respect to DHQS and alters the identity and orientation of a key Arg residue. This and other active site differences in ValA are mostly localized to areas where the ValA substrate differs from that of DHQS. Sequence comparisons with a second SH7PC making a product with distinct stereochemistry lead us to postulate that the product stereochemistry of a given SH7PC is not the result of events taking place during catalysis but is accomplished by selective binding of either the α or β pyranose anomer of the substrate.
Figures



References
-
- Mahmud T. (2003) The C7N aminocyclitol family of natural products. Nat. Prod. Rep. 20, 137–166. - PubMed
-
- Stratmann A.; Mahmud T.; Lee S.; Distler J.; Floss H. G.; Piepersberg W. (1999) The AcbC protein from Actinoplanes species is a C-7-cyclitol synthase related to 3-dehydroquinate synthases and is involved in the biosynthesis of the α-glucosidase inhibitor acarbose. J. Biol. Chem. 274, 10889–10896. - PubMed
-
- Yu Y.; Bai L. Q.; Minagawa K.; Jian X. H.; Li L.; Li J. L.; Chen S. Y.; Cao E. H.; Mahmud T.; Floss H. G.; Zhou X. F.; Deng Z. X. (2005) Gene cluster responsible for validamycin biosynthesis in Streptomyces hygroscopicus subsp jinggangensis 5008. Appl. Environ. Microbiol. 71, 5066–5076. - PMC - PubMed
-
- Carpenter E. P.; Hawkins A. R.; Frost J. W.; Brown K. A. (1998) Structure of dehydroquinate synthase reveals an active site capable of multistep catalysis. Nature 394, 299–302. - PubMed
-
- Nango E.; Kumasaka T.; Hirayama T.; Tanaka N.; Eguchi T. (2008) Structure of 2-deoxy-scyllo-inosose synthase, a key enzyme in the biosynthesis of 2-deoxystreptamine-containing aminoglycoside antibiotics, in complex with a mechanism-based inhibitor and NAD+. Proteins: Struct., Funct., Bioinf. 70, 517–527. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
