

Transcriptional regulation of Caenorhabditis elegans FOXO/DAF-16 modulates lifespan
- PMID: 24834345
- PMCID: PMC4022319
- DOI: 10.1186/2046-2395-3-5
Transcriptional regulation of Caenorhabditis elegans FOXO/DAF-16 modulates lifespan
Abstract
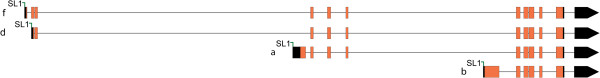
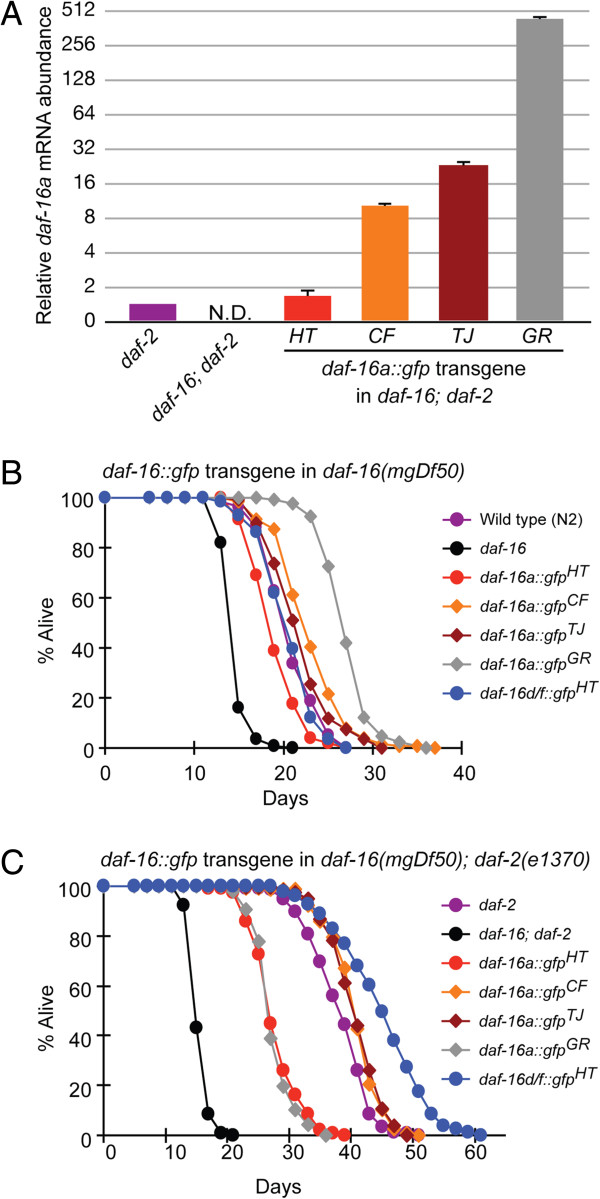
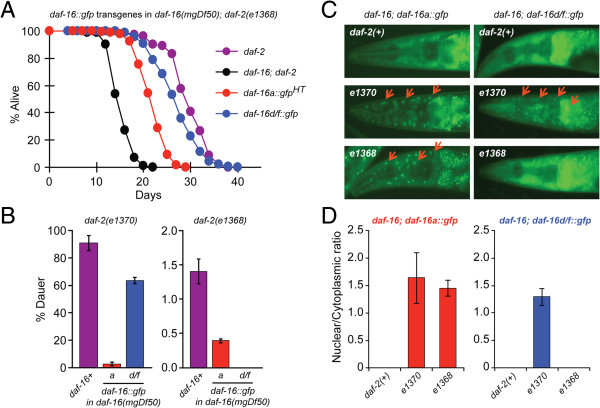
Background: Insulin/IGF-1 signaling plays a central role in longevity across phylogeny. In C. elegans, the forkhead box O (FOXO) transcription factor, DAF-16, is the primary target of insulin/IGF-1 signaling, and multiple isoforms of DAF-16 (a, b, and d/f) modulate lifespan, metabolism, dauer formation, and stress resistance. Thus far, across phylogeny modulation of mammalian FOXOs and DAF-16 have focused on post-translational regulation with little focus on transcriptional regulation. In C. elegans, we have previously shown that DAF-16d/f cooperates with DAF-16a to promote longevity. In this study, we generated transgenic strains expressing near-endogenous levels of either daf-16a or daf-16d/f, and examined temporal expression of the isoforms to further define how these isoforms contribute to lifespan regulation.
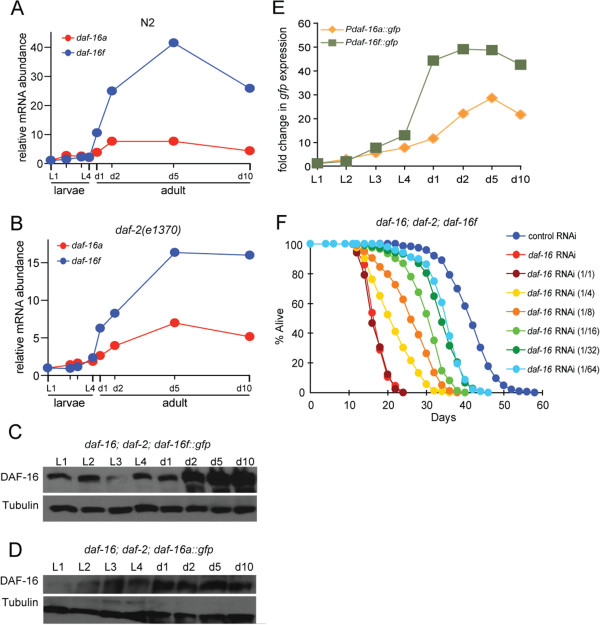
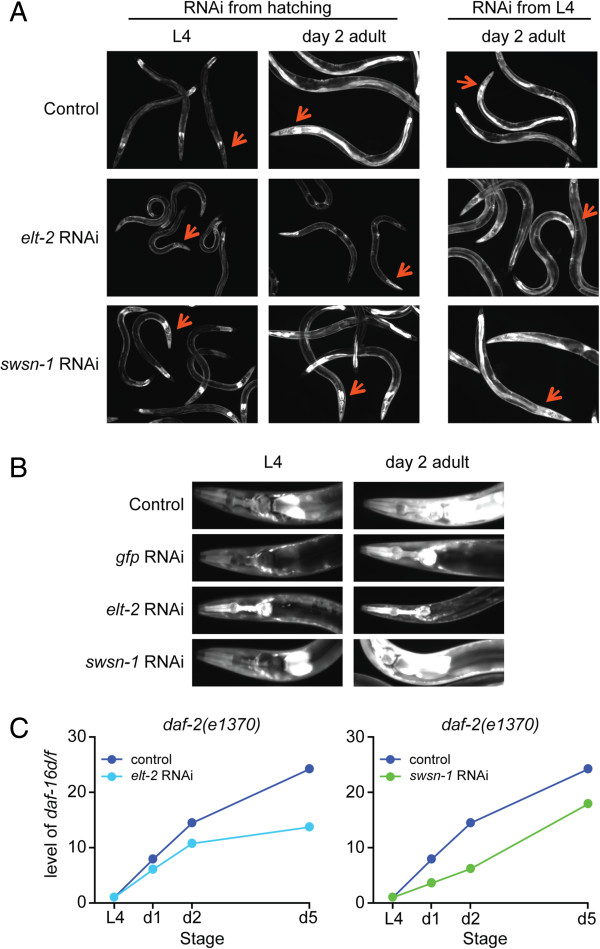
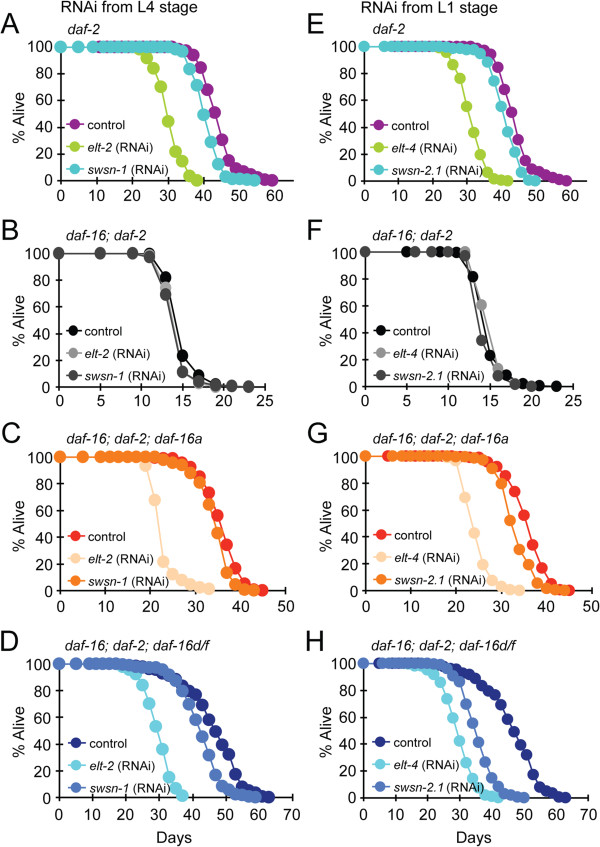
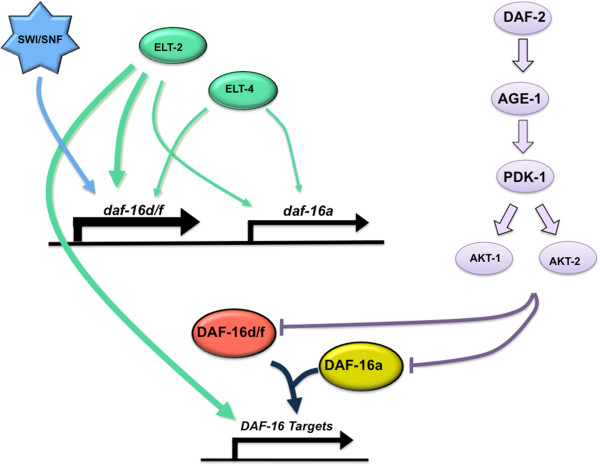
Results: Here, we show that DAF-16a is sensitive both to changes in gene dosage and to alterations in the level of insulin/IGF-1 signaling. Interestingly, we find that as worms age, the intestinal expression of daf-16d/f but not daf-16a is dramatically upregulated at the level of transcription. Preventing this transcriptional upregulation shortens lifespan, indicating that transcriptional regulation of daf-16d/f promotes longevity. In an RNAi screen of transcriptional regulators, we identify elt-2 (GATA transcription factor) and swsn-1 (core subunit of SWI/SNF complex) as key modulators of daf-16d/f gene expression. ELT-2 and another GATA factor, ELT-4, promote longevity via both DAF-16a and DAF-16d/f while the components of SWI/SNF complex promote longevity specifically via DAF-16d/f.
Conclusions: Our findings indicate that transcriptional control of C. elegans FOXO/daf-16 is an essential regulatory event. Considering the conservation of FOXO across species, our findings identify a new layer of FOXO regulation as a potential determinant of mammalian longevity and age-related diseases such as cancer and diabetes.
Keywords: Aging; C. elegans; DAF-16/FOXO; Isoforms; Longevity; Transcription.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
