

Spatiotemporal receptive fields of barrel cortex revealed by reverse correlation of synaptic input
- PMID: 24836076
- PMCID: PMC4203687
- DOI: 10.1038/nn.3720
Spatiotemporal receptive fields of barrel cortex revealed by reverse correlation of synaptic input
Abstract
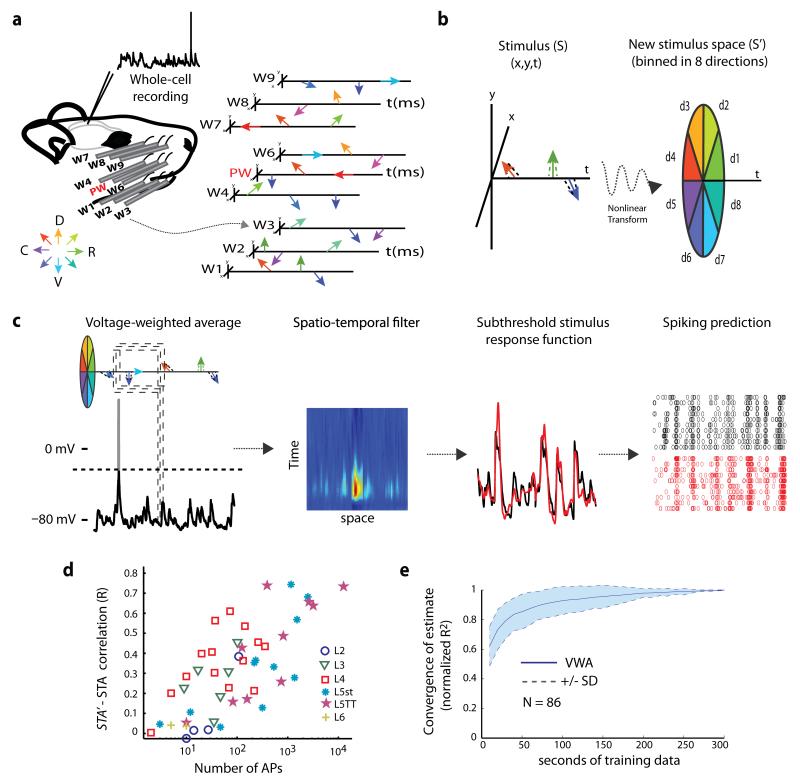
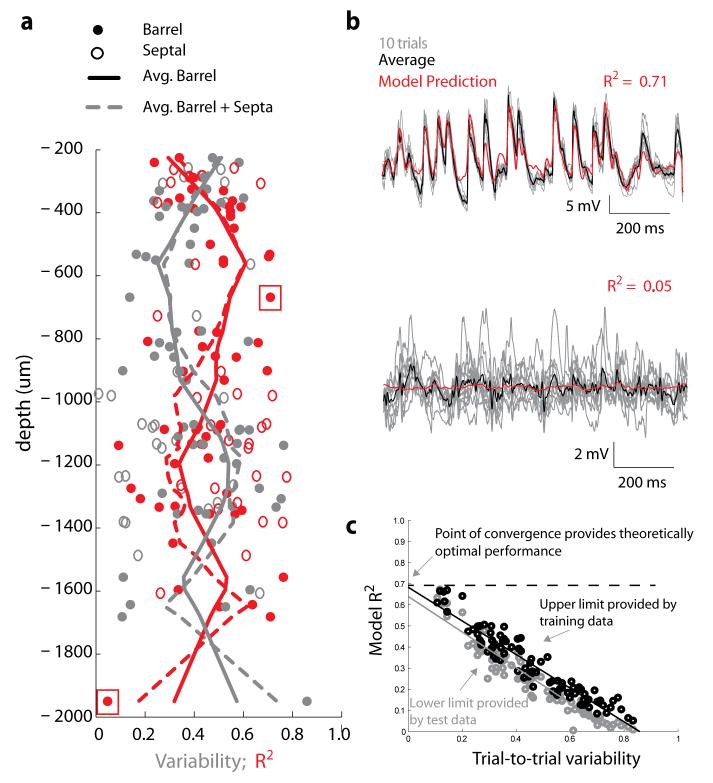
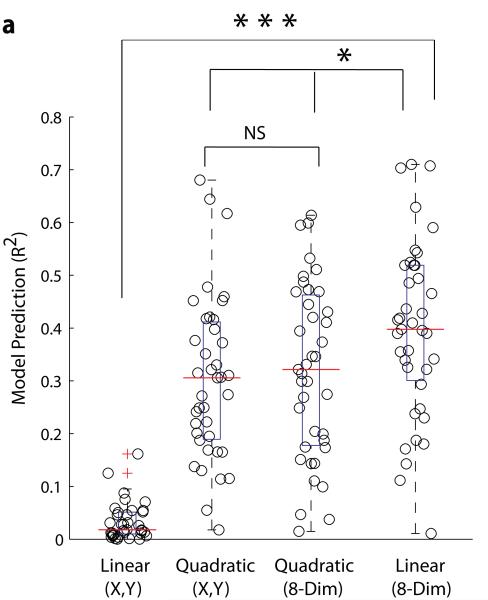
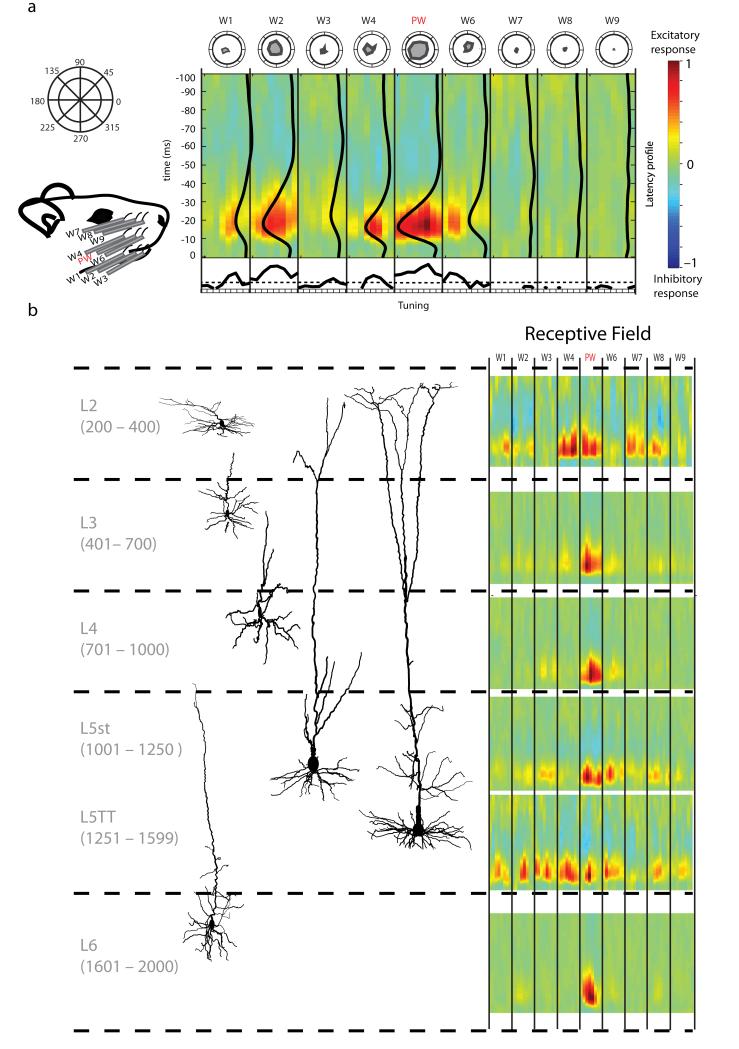
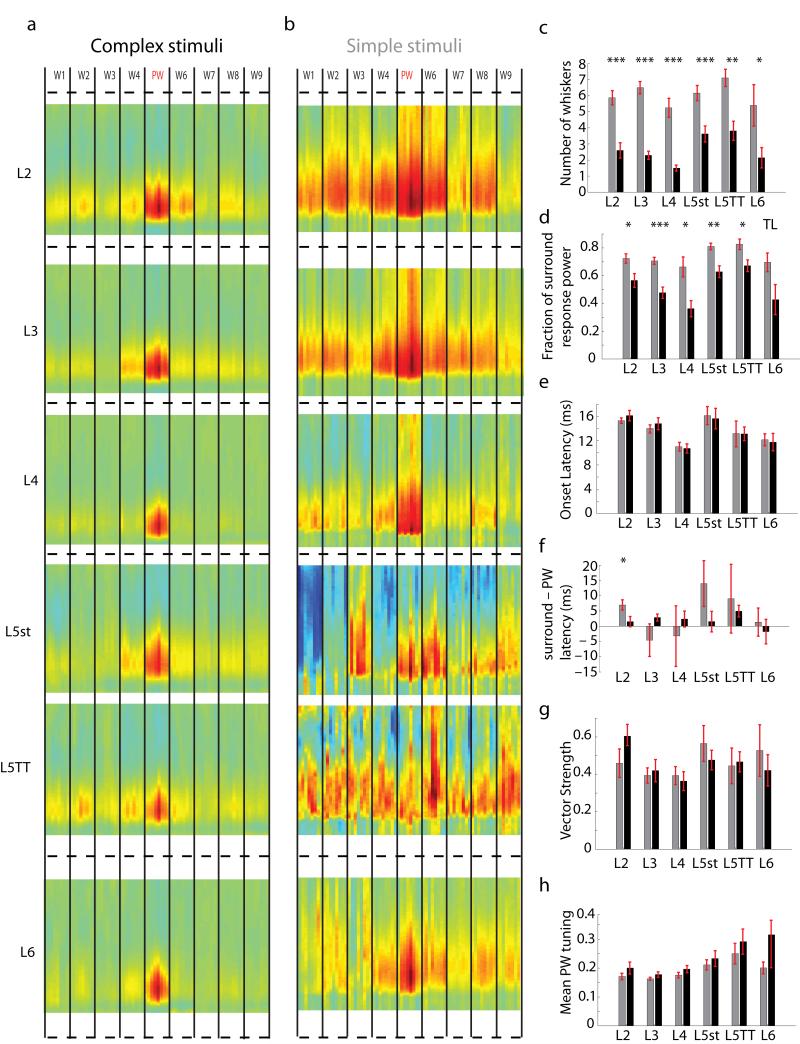
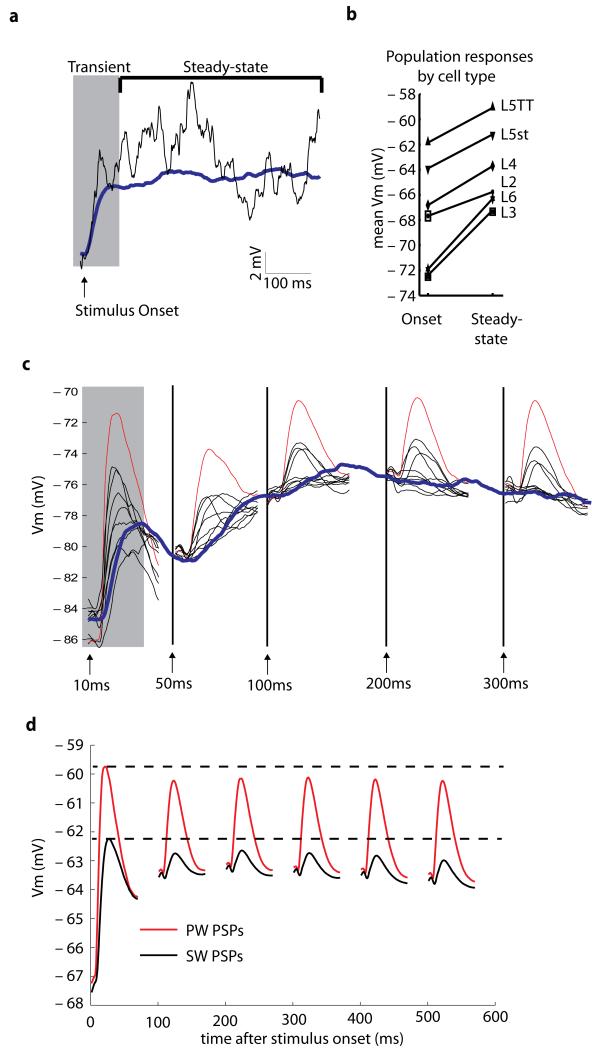
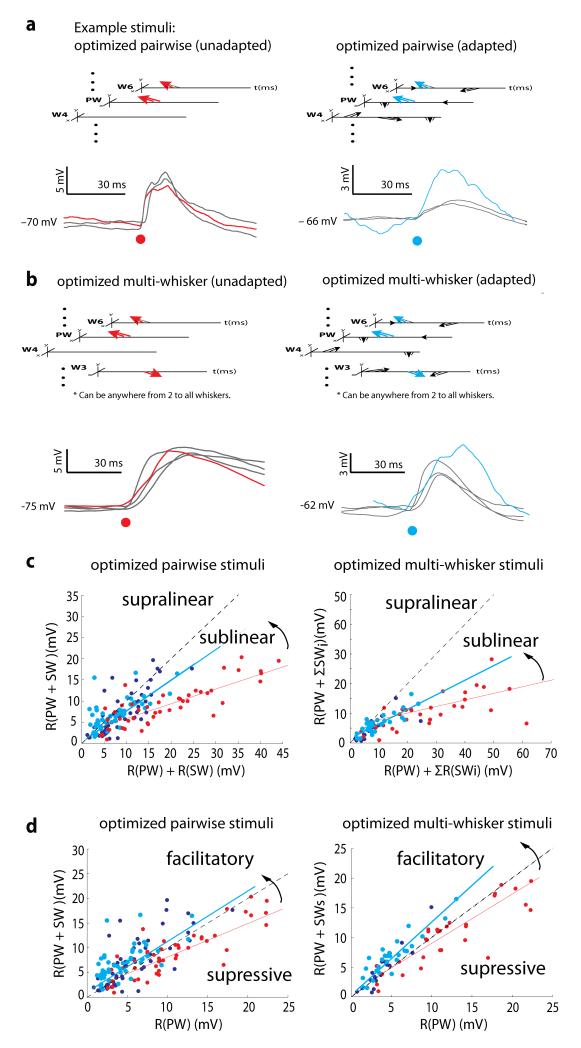
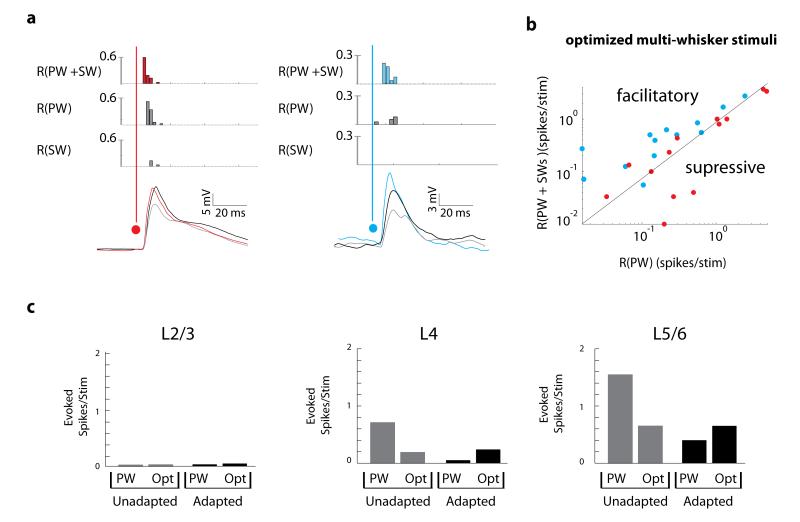
Of all of the sensory areas, barrel cortex is among the best understood in terms of circuitry, yet least understood in terms of sensory function. We combined intracellular recording in rats with a multi-directional, multi-whisker stimulator system to estimate receptive fields by reverse correlation of stimuli to synaptic inputs. Spatiotemporal receptive fields were identified orders of magnitude faster than by conventional spike-based approaches, even for neurons with little spiking activity. Given a suitable stimulus representation, a linear model captured the stimulus-response relationship for all neurons with high accuracy. In contrast with conventional single-whisker stimuli, complex stimuli revealed markedly sharpened receptive fields, largely as a result of adaptation. This phenomenon allowed the surround to facilitate rather than to suppress responses to the principal whisker. Optimized stimuli enhanced firing in layers 4-6, but not in layers 2/3, which remained sparsely active. Surround facilitation through adaptation may be required for discriminating complex shapes and textures during natural sensing.
Figures
Comment in
-
The needle in the haystack.Nat Neurosci. 2014 Jun;17(6):752-3. doi: 10.1038/nn.3730. Nat Neurosci. 2014. PMID: 24866038 No abstract available.
References
-
- Moore CI, Nelson SB. Spatio-temporal subthreshold receptive fields in the vibrissa representation of rat primary somatosensory cortex. J Neurophysiol. 1998;80:2882–2892. - PubMed
-
- Zhu JJ, Connors BW. Intrinsic firing patterns and whisker-evoked synaptic responses of neurons in the rat barrel cortex. J Neurophysiol. 1999;81:1171–1183. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
