

Elevated sphingosine-1-phosphate promotes sickling and sickle cell disease progression
- PMID: 24837436
- PMCID: PMC4089467
- DOI: 10.1172/JCI74604
Elevated sphingosine-1-phosphate promotes sickling and sickle cell disease progression
Abstract
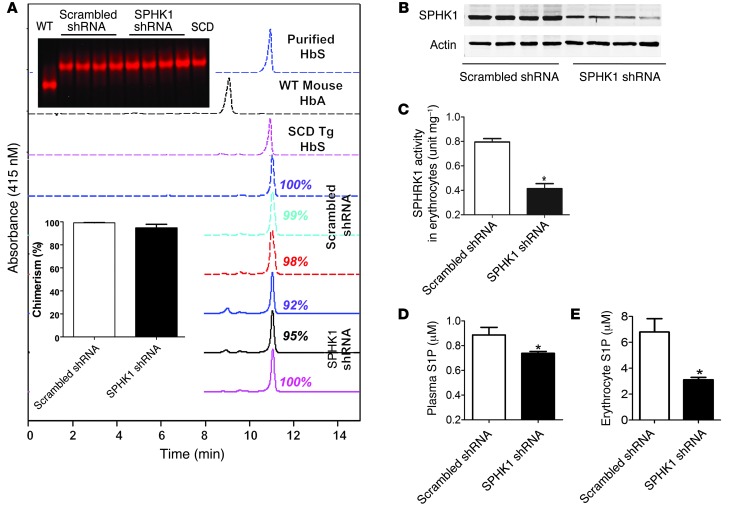
Sphingosine-1-phosphate (S1P) is a bioactive lipid that regulates multicellular functions through interactions with its receptors on cell surfaces. S1P is enriched and stored in erythrocytes; however, it is not clear whether alterations in S1P are involved in the prevalent and debilitating hemolytic disorder sickle cell disease (SCD). Here, using metabolomic screening, we found that S1P is highly elevated in the blood of mice and humans with SCD. In murine models of SCD, we demonstrated that elevated erythrocyte sphingosine kinase 1 (SPHK1) underlies sickling and disease progression by increasing S1P levels in the blood. Additionally, we observed elevated SPHK1 activity in erythrocytes and increased S1P in blood collected from patients with SCD and demonstrated a direct impact of elevated SPHK1-mediated production of S1P on sickling that was independent of S1P receptor activation in isolated erythrocytes. Together, our findings provide insights into erythrocyte pathophysiology, revealing that a SPHK1-mediated elevation of S1P contributes to sickling and promotes disease progression, and highlight potential therapeutic opportunities for SCD.
Figures







References
-
- Madigan C, Malik P. Pathophysiology and therapy for haemoglobinopathies. Part I: sickle cell disease. Expert Rev Mol Med. 2006;8(9):1–23. - PubMed
-
- Urbinati F, Madigan C, Malik P. Pathophysiology and therapy for haemoglobinopathies. Part II: thalassaemias. Expert Rev Mol Med. 2006;8(10):1–26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
