

Wnt signaling controls the specification of definitive and primitive hematopoiesis from human pluripotent stem cells
- PMID: 24837661
- PMCID: PMC4152856
- DOI: 10.1038/nbt.2915
Wnt signaling controls the specification of definitive and primitive hematopoiesis from human pluripotent stem cells
Abstract
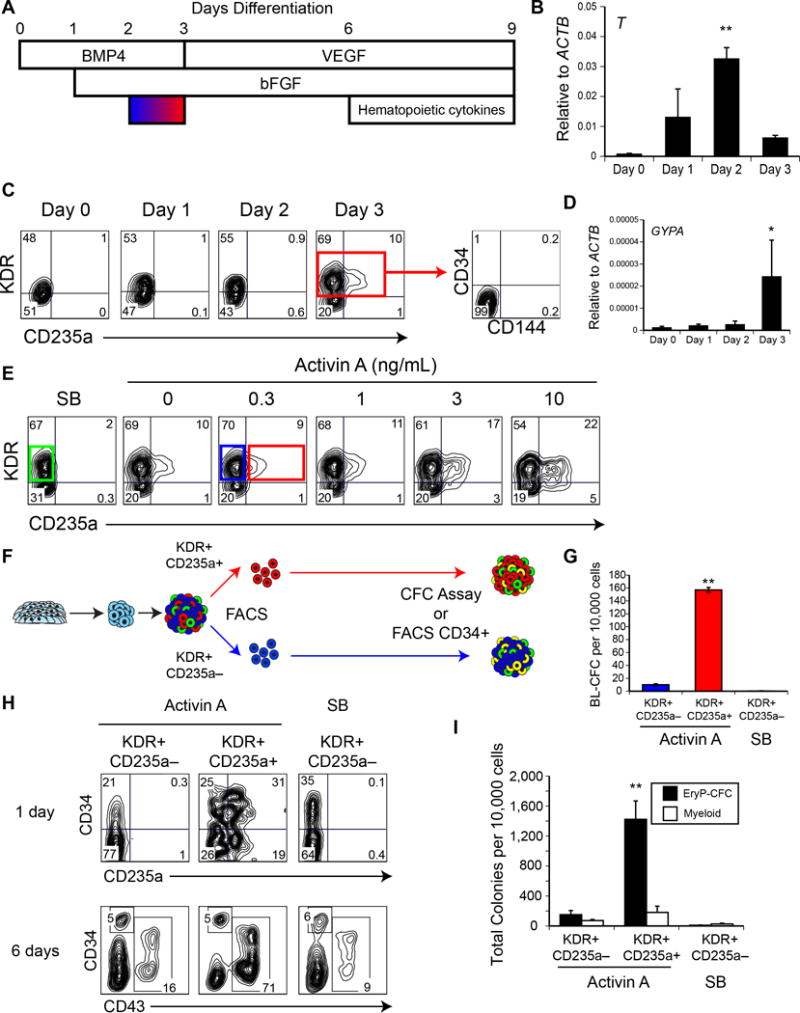
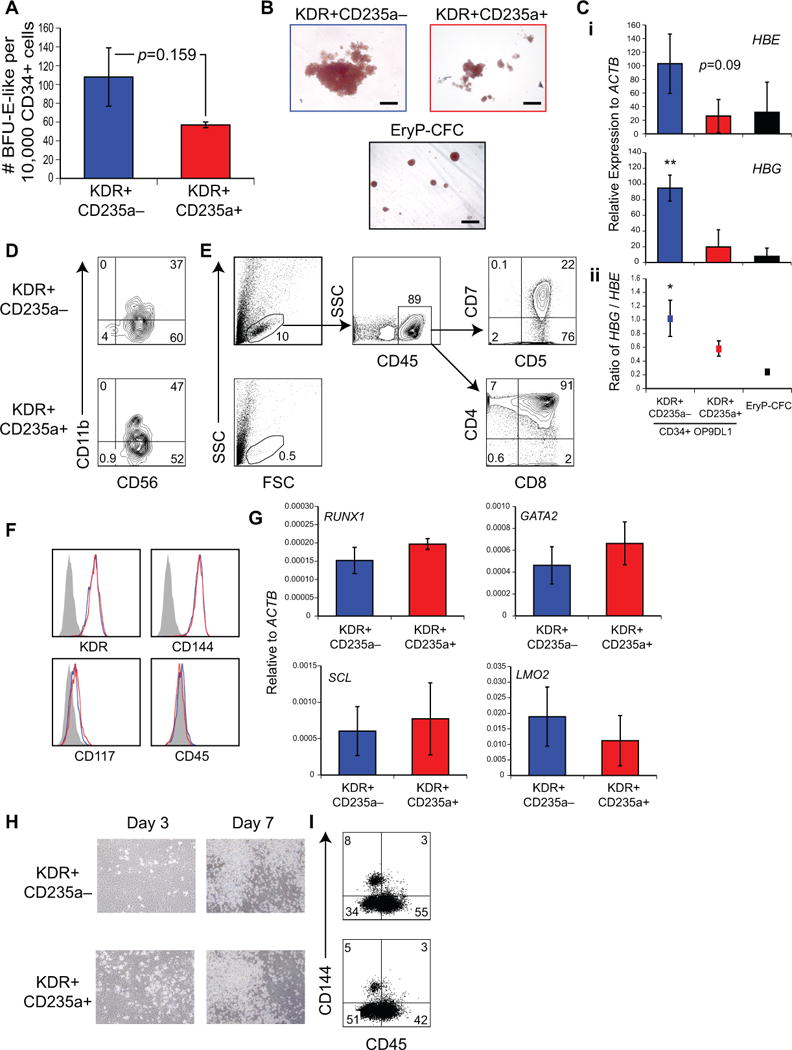
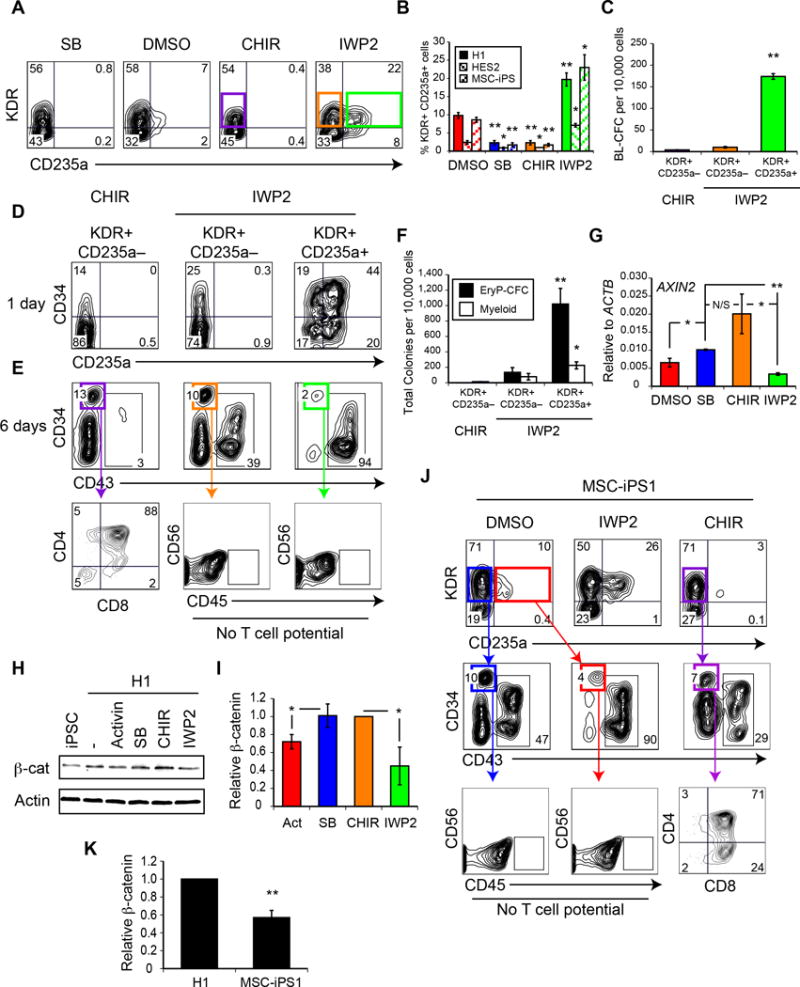
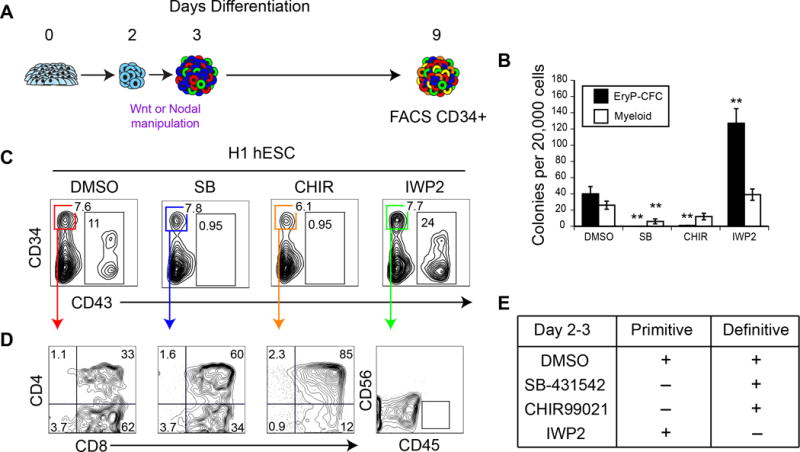
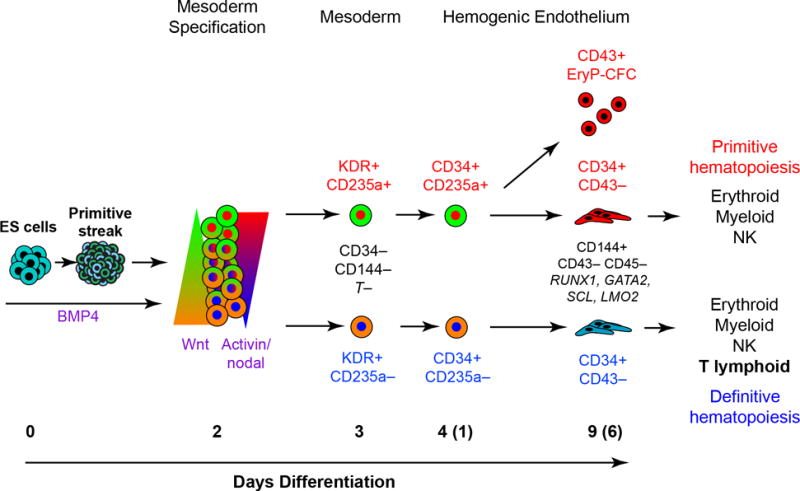
Efforts to derive hematopoietic stem cells (HSCs) from human pluripotent stem cells (hPSCs) are complicated by the fact that embryonic hematopoiesis consists of two programs, primitive and definitive, that differ in developmental potential. As only definitive hematopoiesis generates HSCs, understanding how this program develops is essential for being able to produce this cell population in vitro. Here we show that both hematopoietic programs transition through hemogenic endothelial intermediates and develop from KDR(+)CD34(-)CD144(-) progenitors that are distinguished by CD235a expression. Generation of primitive progenitors (KDR(+)CD235a(+)) depends on stage-specific activin-nodal signaling and inhibition of the Wnt-β-catenin pathway, whereas specification of definitive progenitors (KDR(+)CD235a(-)) requires Wnt-β-catenin signaling during this same time frame. Together, these findings establish simple selective differentiation strategies for the generation of primitive or definitive hematopoietic progenitors by Wnt-β-catenin manipulation, and in doing so provide access to enriched populations for future studies on hPSC-derived hematopoietic development.
Conflict of interest statement
Figures
Comment in
-
Inducing definitive hematopoiesis in a dish.Nat Biotechnol. 2014 Jun;32(6):539-41. doi: 10.1038/nbt.2929. Nat Biotechnol. 2014. PMID: 24911496 No abstract available.
References
-
- Medvinsky A, Rybtsov S, Taoudi S. Embryonic origin of the adult hematopoietic system: advances and questions. Development (Cambridge, England) 2011;138:1017–1031. - PubMed
-
- Huber TL, Kouskoff V, Fehling HJ, Palis J, Keller G. Haemangioblast commitment is initiated in the primitive streak of the mouse embryo. Nature. 2004;432:625–630. - PubMed
-
- Antas VI, Al-Drees MA, Prudence AJ, Sugiyama D, Fraser ST. Hemogenic endothelium: a vessel for blood production. The international journal of biochemistry & cell biology. 2013;45:692–695. - PubMed
-
- Murry CE, Keller G. Differentiation of embryonic stem cells to clinically relevant populations: lessons from embryonic development. Cell. 2008;132:661–680. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
