

RNA Bind-n-Seq: quantitative assessment of the sequence and structural binding specificity of RNA binding proteins
- PMID: 24837674
- PMCID: PMC4142047
- DOI: 10.1016/j.molcel.2014.04.016
RNA Bind-n-Seq: quantitative assessment of the sequence and structural binding specificity of RNA binding proteins
Abstract
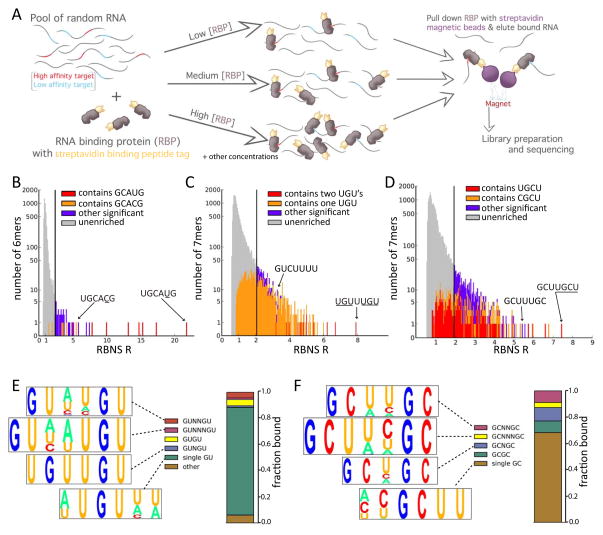
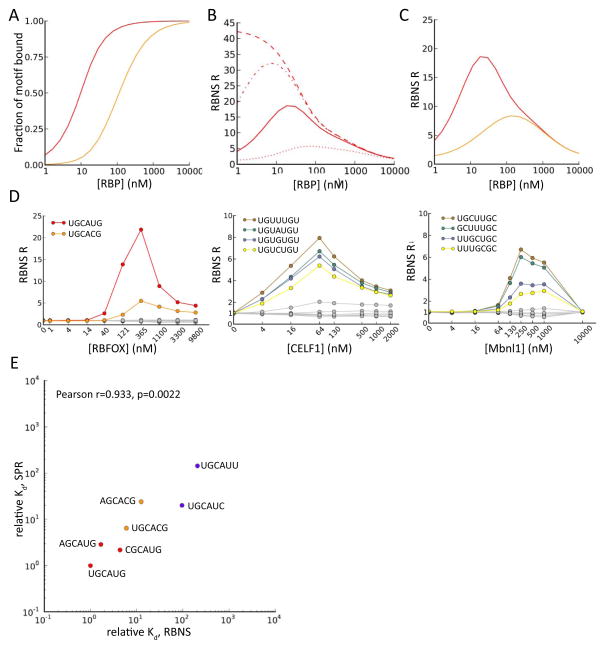
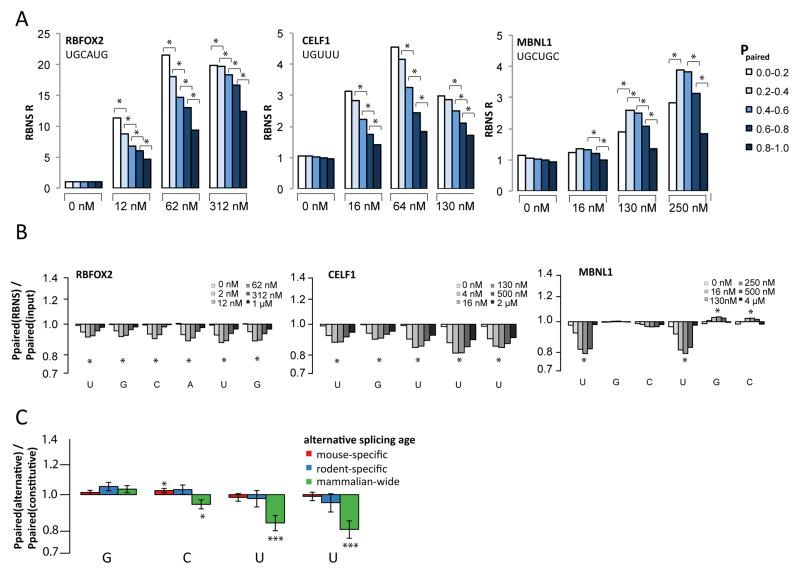
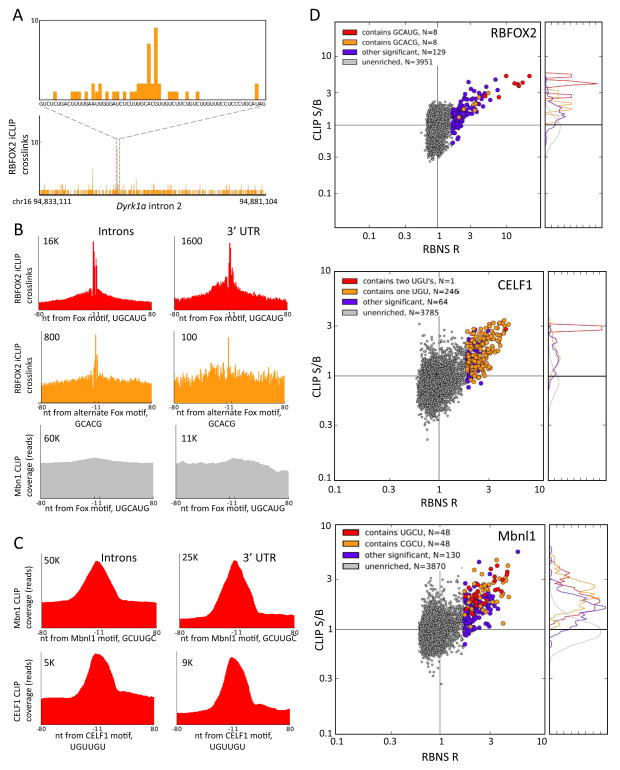
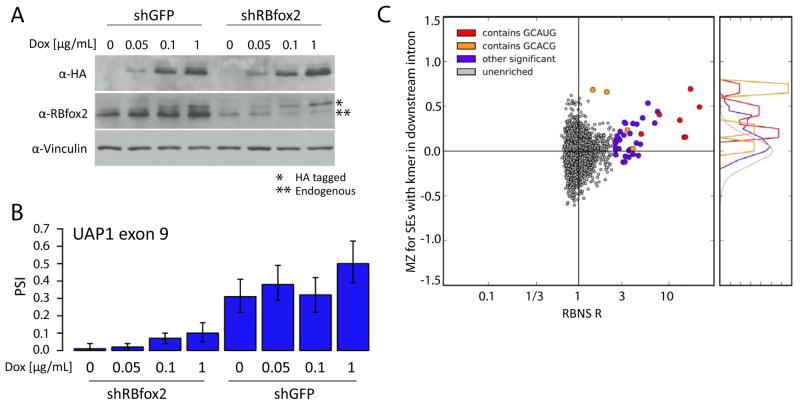
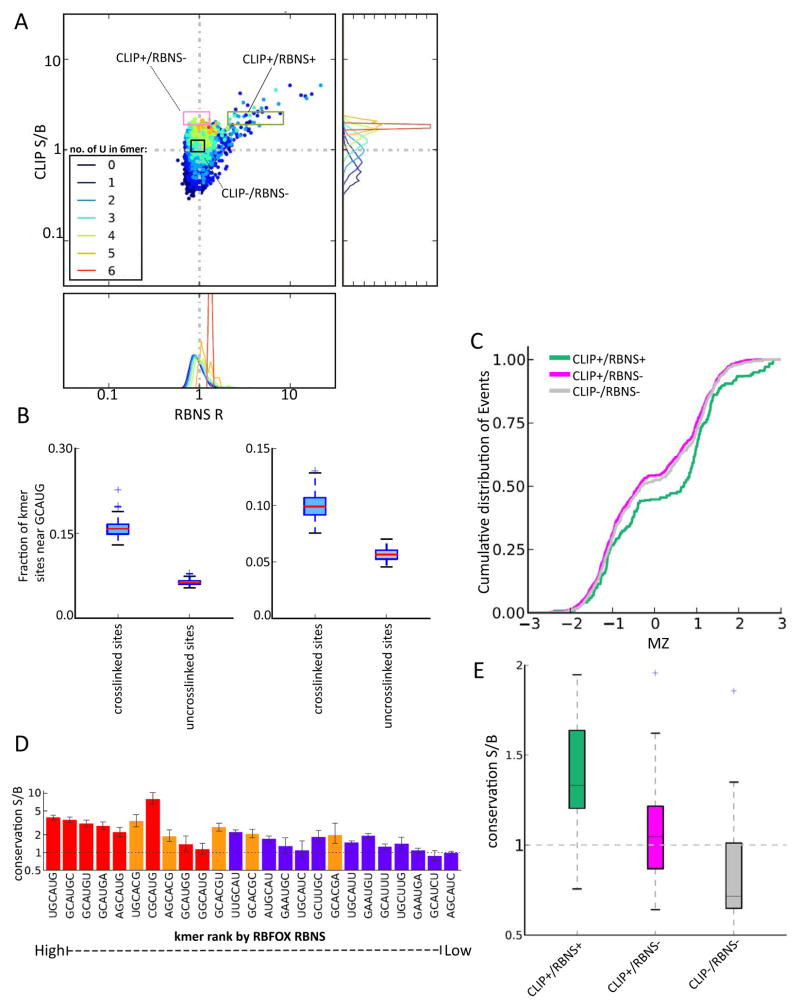
Specific protein-RNA interactions guide posttranscriptional gene regulation. Here, we describe RNA Bind-n-Seq (RBNS), a method that comprehensively characterizes sequence and structural specificity of RNA binding proteins (RBPs), and its application to the developmental alternative splicing factors RBFOX2, CELF1/CUGBP1, and MBNL1. For each factor, we recovered both canonical motifs and additional near-optimal binding motifs. RNA secondary structure inhibits binding of RBFOX2 and CELF1, while MBNL1 favors unpaired Us but tolerates C/G pairing in motifs containing UGC and/or GCU. Dissociation constants calculated from RBNS data using a novel algorithm correlated highly with values measured by surface plasmon resonance. Motifs identified by RBNS were conserved, were bound and active in vivo, and distinguished the subset of motifs enriched by CLIP-Seq that had regulatory activity. Together, our data demonstrate that RBNS complements crosslinking-based methods and show that in vivo binding and activity of these splicing factors is driven largely by intrinsic RNA affinity.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
