

Partitioning the metabolic cost of human running: a task-by-task approach
- PMID: 24838747
- PMCID: PMC4296200
- DOI: 10.1093/icb/icu033
Partitioning the metabolic cost of human running: a task-by-task approach
Erratum in
-
Erratum.Integr Comp Biol. 2017 Jul 1;57(1):169. doi: 10.1093/icb/icx003. Integr Comp Biol. 2017. PMID: 28369435 Free PMC article. No abstract available.
Abstract
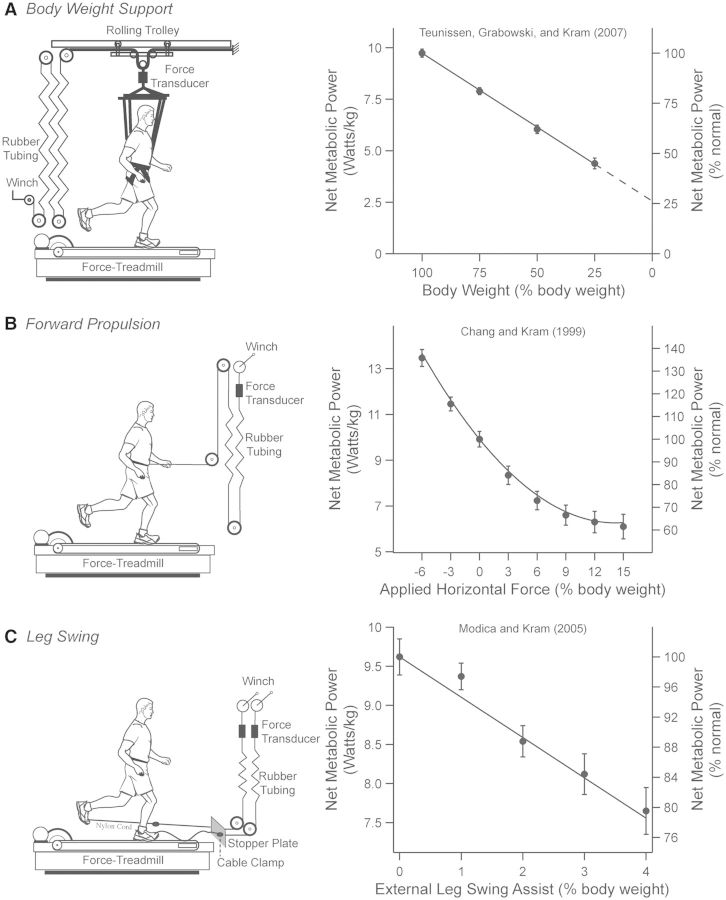
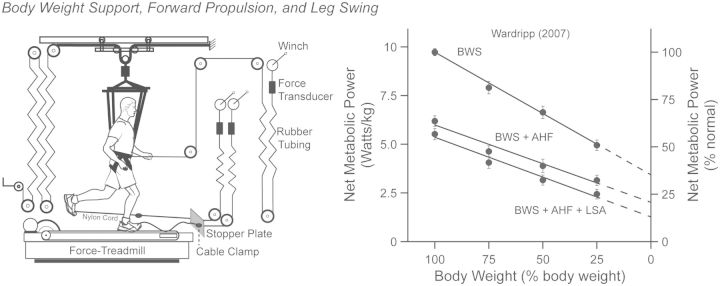
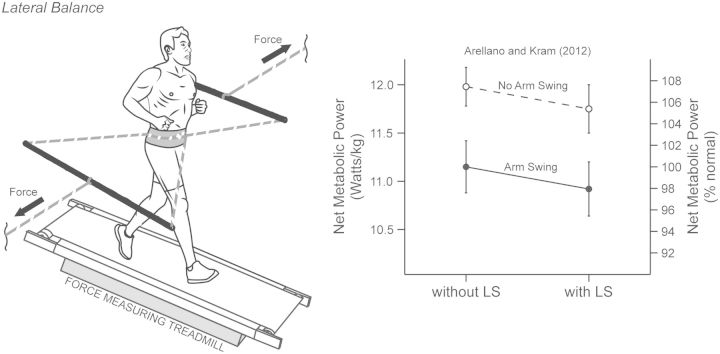
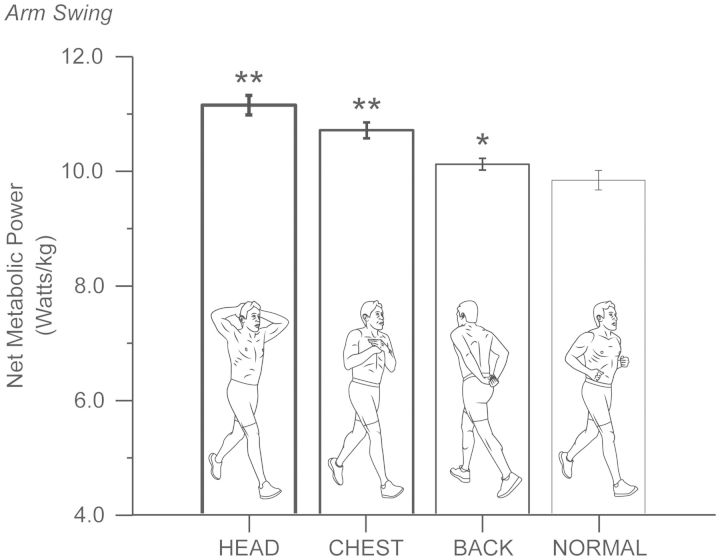
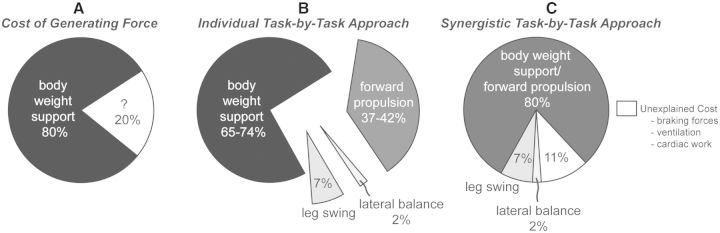
Compared with other species, humans can be very tractable and thus an ideal "model system" for investigating the metabolic cost of locomotion. Here, we review the biomechanical basis for the metabolic cost of running. Running has been historically modeled as a simple spring-mass system whereby the leg acts as a linear spring, storing, and returning elastic potential energy during stance. However, if running can be modeled as a simple spring-mass system with the underlying assumption of perfect elastic energy storage and return, why does running incur a metabolic cost at all? In 1980, Taylor et al. proposed the "cost of generating force" hypothesis, which was based on the idea that elastic structures allow the muscles to transform metabolic energy into force, and not necessarily mechanical work. In 1990, Kram and Taylor then provided a more explicit and quantitative explanation by demonstrating that the rate of metabolic energy consumption is proportional to body weight and inversely proportional to the time of foot-ground contact for a variety of animals ranging in size and running speed. With a focus on humans, Kram and his colleagues then adopted a task-by-task approach and initially found that the metabolic cost of running could be "individually" partitioned into body weight support (74%), propulsion (37%), and leg-swing (20%). Summing all these biomechanical tasks leads to a paradoxical overestimation of 131%. To further elucidate the possible interactions between these tasks, later studies quantified the reductions in metabolic cost in response to synergistic combinations of body weight support, aiding horizontal forces, and leg-swing-assist forces. This synergistic approach revealed that the interactive nature of body weight support and forward propulsion comprises ∼80% of the net metabolic cost of running. The task of leg-swing at most comprises ∼7% of the net metabolic cost of running and is independent of body weight support and forward propulsion. In our recent experiments, we have continued to refine this task-by-task approach, demonstrating that maintaining lateral balance comprises only 2% of the net metabolic cost of running. In contrast, arm-swing reduces the cost by ∼3%, indicating a net metabolic benefit. Thus, by considering the synergistic nature of body weight support and forward propulsion, as well as the tasks of leg-swing and lateral balance, we can account for 89% of the net metabolic cost of human running.
© The Author 2014. Published by Oxford University Press on behalf of the Society for Integrative and Comparative Biology. All rights reserved. For permissions please email: journals.permissions@oup.com.
Figures
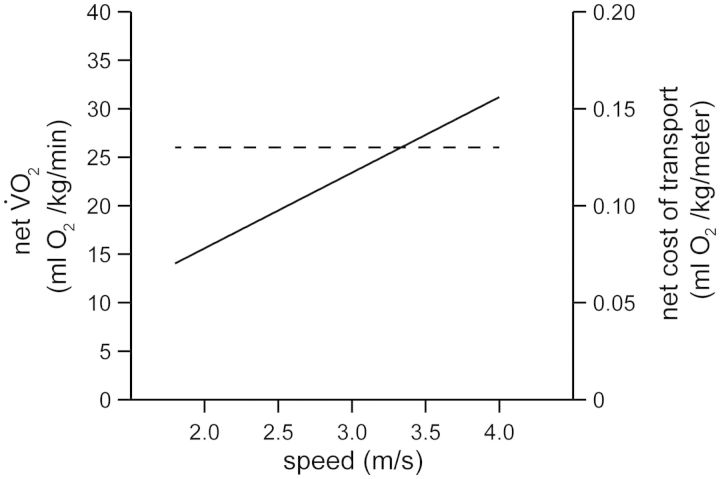
 and running speed crossed the zero-speed intercept at a value that was similar to his subjects’ resting metabolism. After subtracting this value from gross
and running speed crossed the zero-speed intercept at a value that was similar to his subjects’ resting metabolism. After subtracting this value from gross  , Margaria et al. (1963) referred to this quantity using the term “net”.
, Margaria et al. (1963) referred to this quantity using the term “net”.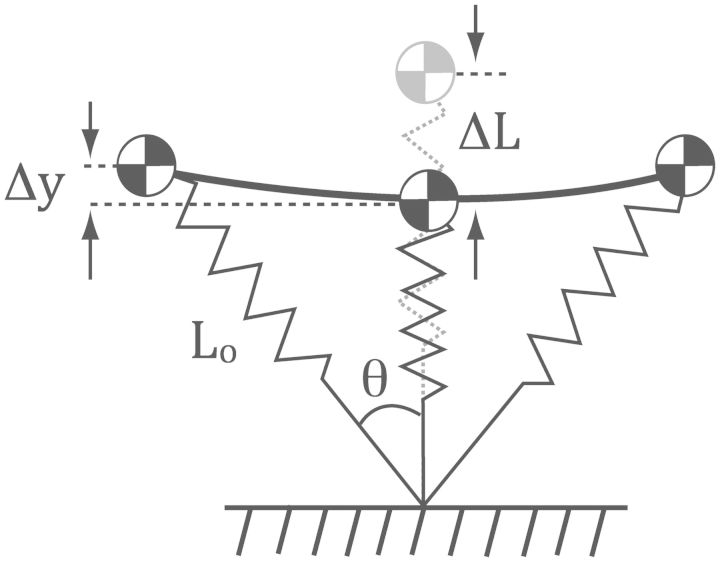
References
-
- Aaron EA, Johnson BD, Seow CK, Dempsey JA. Oxygen cost of exercise hyperpnea: measurement. J Appl Physiol. 1992a;72:1810–7. - PubMed
-
- Aaron EA, Seow KC, Johnson BD, Dempsey JA. Oxygen cost of exercise hyperpnea: implications for performance. J Appl Physiol. 1992b;72:1818–25. - PubMed
-
- Alexander RM. Elastic energy stores in running vertebraties. Am Zool. 1984;24:85–94.
-
- Alexander RM, Bennetclark HC. Storage of elastic strain-energy in muscle and other tissues. Nature. 1977;265:114–7. - PubMed
-
- Arellano CJ, Kram R. The effects of step width and arm swing on energetic cost and lateral balance during running. J Biomech. 2011;44:1291–5. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
