

A causal relation between bioluminescence and oxygen to quantify the cell niche
- PMID: 24840204
- PMCID: PMC4026314
- DOI: 10.1371/journal.pone.0097572
A causal relation between bioluminescence and oxygen to quantify the cell niche
Erratum in
-
Correction: A Causal Relation between Bioluminescence and Oxygen to Quantify the Cell Niche.PLoS One. 2015 May 28;10(5):e0128852. doi: 10.1371/journal.pone.0128852. eCollection 2015. PLoS One. 2015. PMID: 26020777 Free PMC article. No abstract available.
Abstract
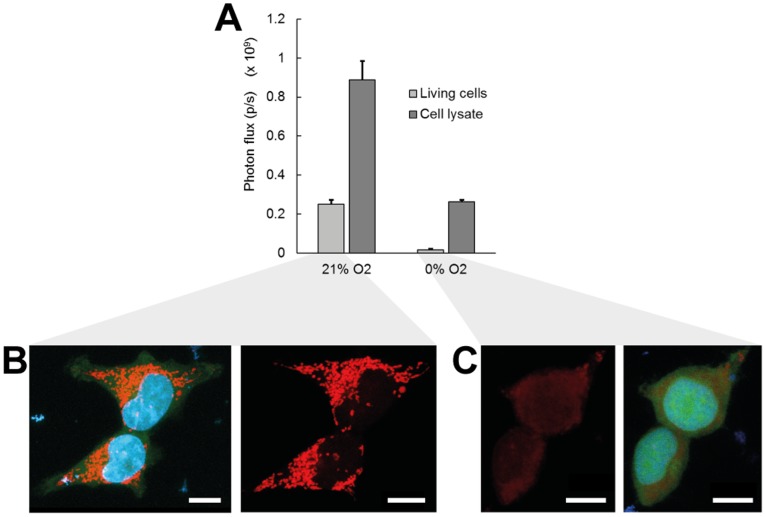
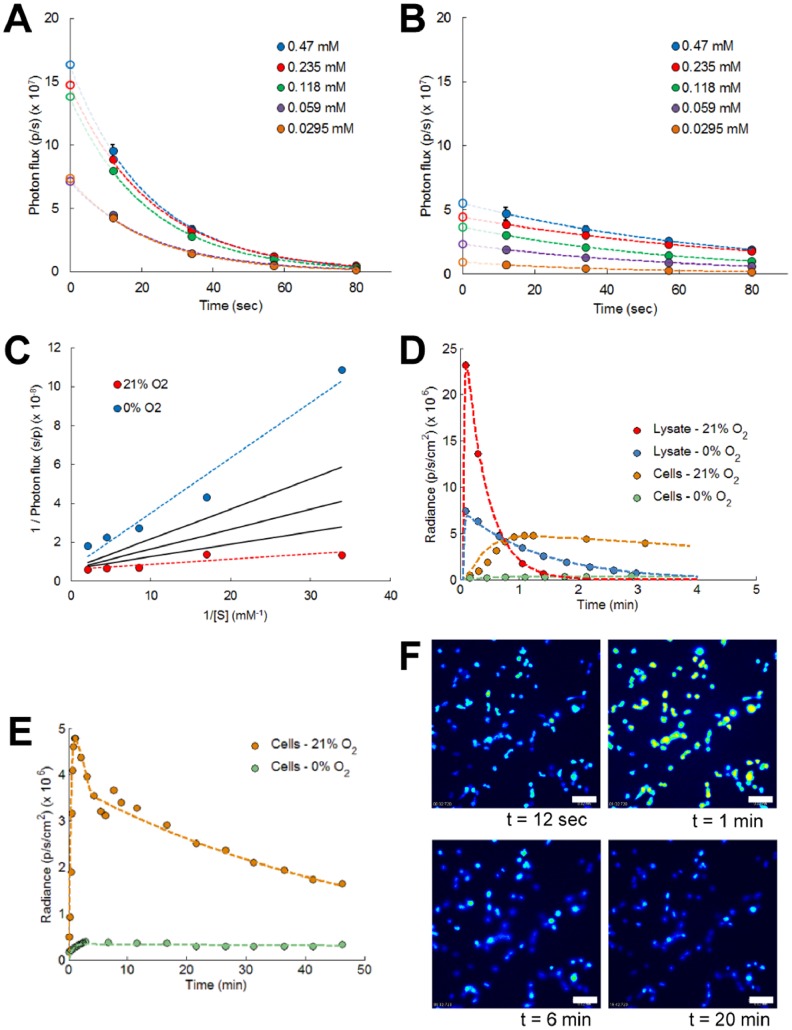
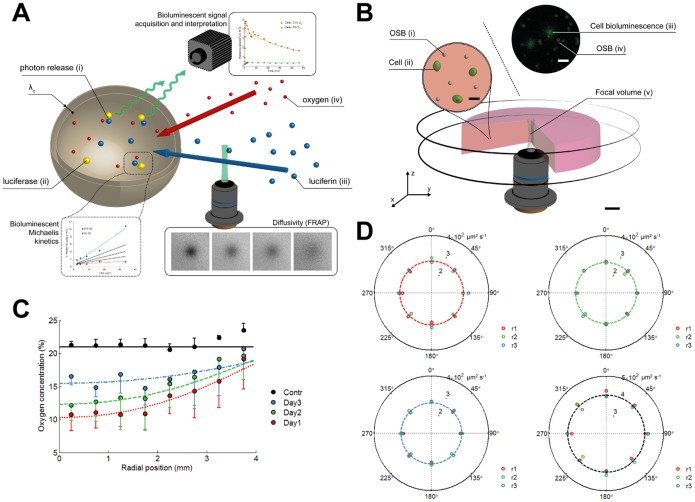
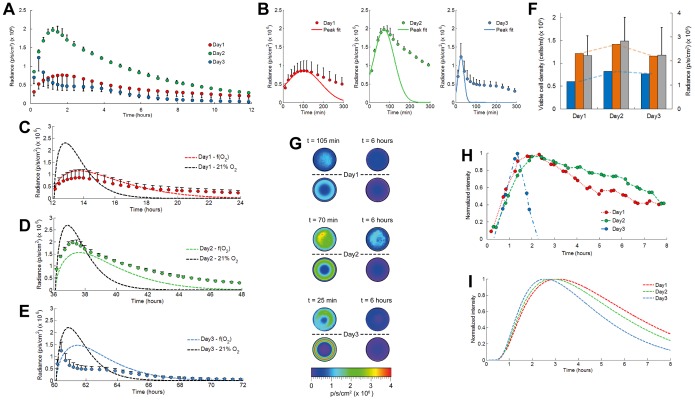
Bioluminescence imaging assays have become a widely integrated technique to quantify effectiveness of cell-based therapies by monitoring fate and survival of transplanted cells. To date these assays are still largely qualitative and often erroneous due to the complexity and dynamics of local micro-environments (niches) in which the cells reside. Here, we report, using a combined experimental and computational approach, on oxygen that besides being a critical niche component responsible for cellular energy metabolism and cell-fate commitment, also serves a primary role in regulating bioluminescent light kinetics. We demonstrate the potential of an oxygen dependent Michaelis-Menten relation in quantifying intrinsic bioluminescence intensities by resolving cell-associated oxygen gradients from bioluminescent light that is emitted from three-dimensional (3D) cell-seeded hydrogels. Furthermore, the experimental and computational data indicate a strong causal relation of oxygen concentration with emitted bioluminescence intensities. Altogether our approach demonstrates the importance of oxygen to evolve towards quantitative bioluminescence and holds great potential for future microscale measurement of oxygen tension in an easily accessible manner.
Conflict of interest statement
Figures
Similar articles
-
Reporter cell activity within hydrogel constructs quantified from oxygen-independent bioluminescence.Biomaterials. 2014 Sep;35(28):8065-77. doi: 10.1016/j.biomaterials.2014.06.002. Epub 2014 Jun 21. Biomaterials. 2014. PMID: 24957291
-
High-Throughput Bioluminescence Imaging and Reporter Gene Assay with 3D Spheroids from Human Cell Lines.Methods Mol Biol. 2020;2081:3-14. doi: 10.1007/978-1-4939-9940-8_1. Methods Mol Biol. 2020. PMID: 31721114
-
Measurement of Bacterial Bioluminescence Intensity and Spectrum: Current Physical Techniques and Principles.Adv Biochem Eng Biotechnol. 2016;154:19-45. doi: 10.1007/10_2015_324. Adv Biochem Eng Biotechnol. 2016. PMID: 25981856 Review.
-
Towards Mimicking the Fetal Liver Niche: The Influence of Elasticity and Oxygen Tension on Hematopoietic Stem/Progenitor Cells Cultured in 3D Fibrin Hydrogels.Int J Mol Sci. 2020 Sep 2;21(17):6367. doi: 10.3390/ijms21176367. Int J Mol Sci. 2020. PMID: 32887387 Free PMC article.
-
The origins of marine bioluminescence: turning oxygen defence mechanisms into deep-sea communication tools.J Exp Biol. 1998 Apr;201(Pt 8):1211-21. doi: 10.1242/jeb.201.8.1211. J Exp Biol. 1998. PMID: 9510532 Review.
Cited by
-
Targeted intracellular delivery of molecular cargo to hypoxic human breast cancer stem cells.bioRxiv [Preprint]. 2024 Nov 12:2024.01.12.575071. doi: 10.1101/2024.01.12.575071. bioRxiv. 2024. Update in: Cancer Commun (Lond). 2025 Jun;45(6):714-718. doi: 10.1002/cac2.70018. PMID: 39605477 Free PMC article. Updated. Preprint.
-
Correction: A Causal Relation between Bioluminescence and Oxygen to Quantify the Cell Niche.PLoS One. 2015 May 28;10(5):e0128852. doi: 10.1371/journal.pone.0128852. eCollection 2015. PLoS One. 2015. PMID: 26020777 Free PMC article. No abstract available.
-
Modular platform for low-light microscopy.Biomed Opt Express. 2015 Oct 27;6(11):4585-98. doi: 10.1364/BOE.6.004585. eCollection 2015 Nov 1. Biomed Opt Express. 2015. PMID: 26601020 Free PMC article.
-
Practical Guidance for Developing Small-Molecule Optical Probes for In Vivo Imaging.Mol Imaging Biol. 2023 Feb;25(1):240-264. doi: 10.1007/s11307-023-01800-1. Epub 2023 Feb 6. Mol Imaging Biol. 2023. PMID: 36745354 Review.
-
A New Perspective on Stroke Research: Unraveling the Role of Brain Oxygen Dynamics in Stroke Pathophysiology.Aging Dis. 2024 Aug 2;16(4):2343-2353. doi: 10.14336/AD.2024.0548. Aging Dis. 2024. PMID: 39226161 Free PMC article. Review.
References
-
- Roda A, Guardigli M, Michelini E, Mirasoli M (2009) Bioluminescence in analytical chemistry and in vivo imaging. Trends Anal Chem 28: 307–22.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
