

The injectable-only contraceptive medroxyprogesterone acetate, unlike norethisterone acetate and progesterone, regulates inflammatory genes in endocervical cells via the glucocorticoid receptor
- PMID: 24840644
- PMCID: PMC4026143
- DOI: 10.1371/journal.pone.0096497
The injectable-only contraceptive medroxyprogesterone acetate, unlike norethisterone acetate and progesterone, regulates inflammatory genes in endocervical cells via the glucocorticoid receptor
Abstract
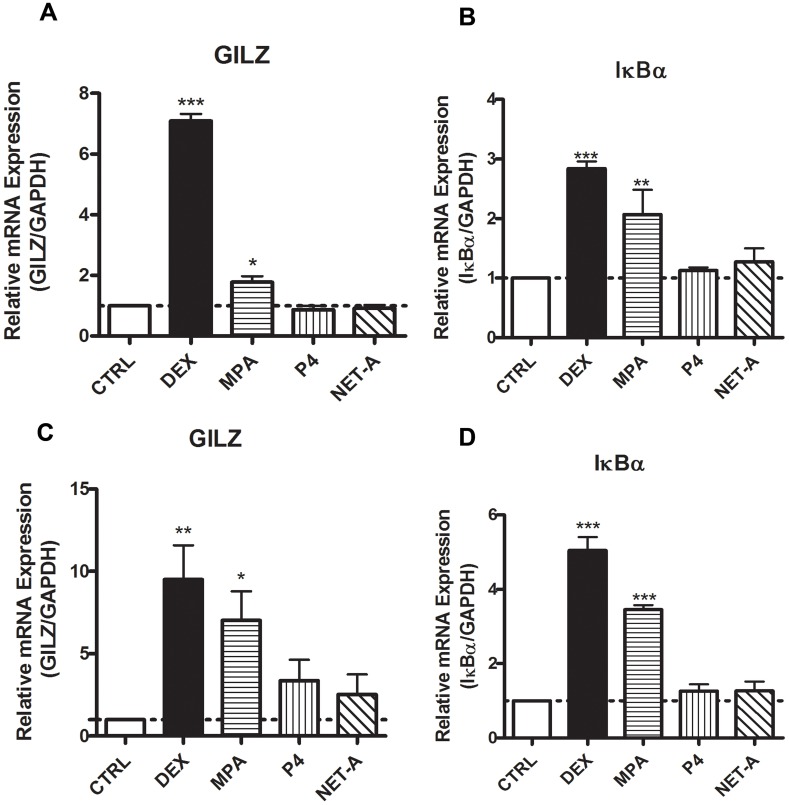
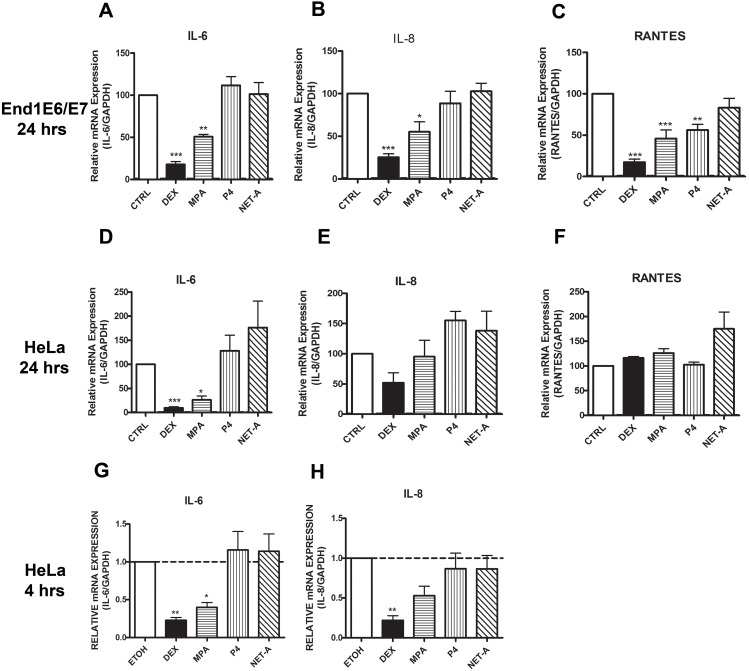
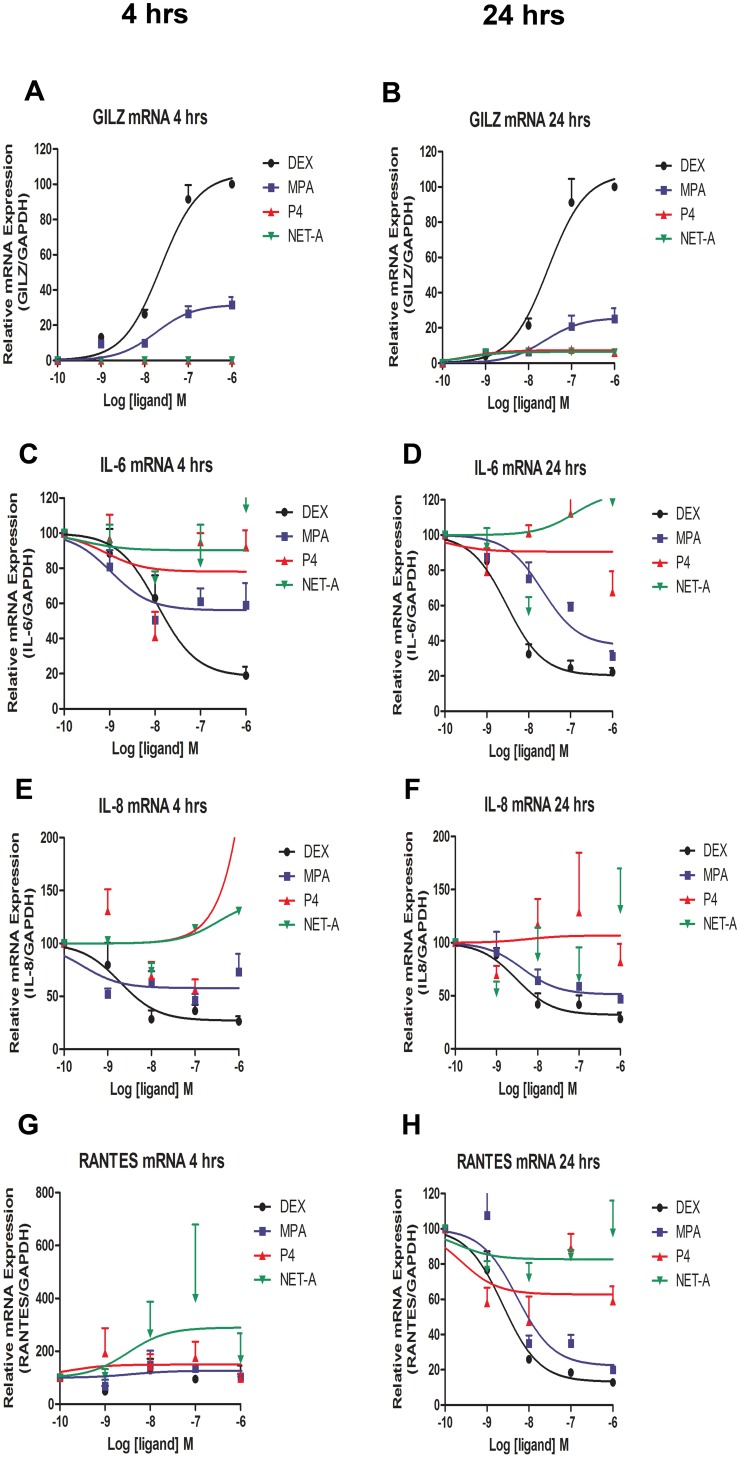
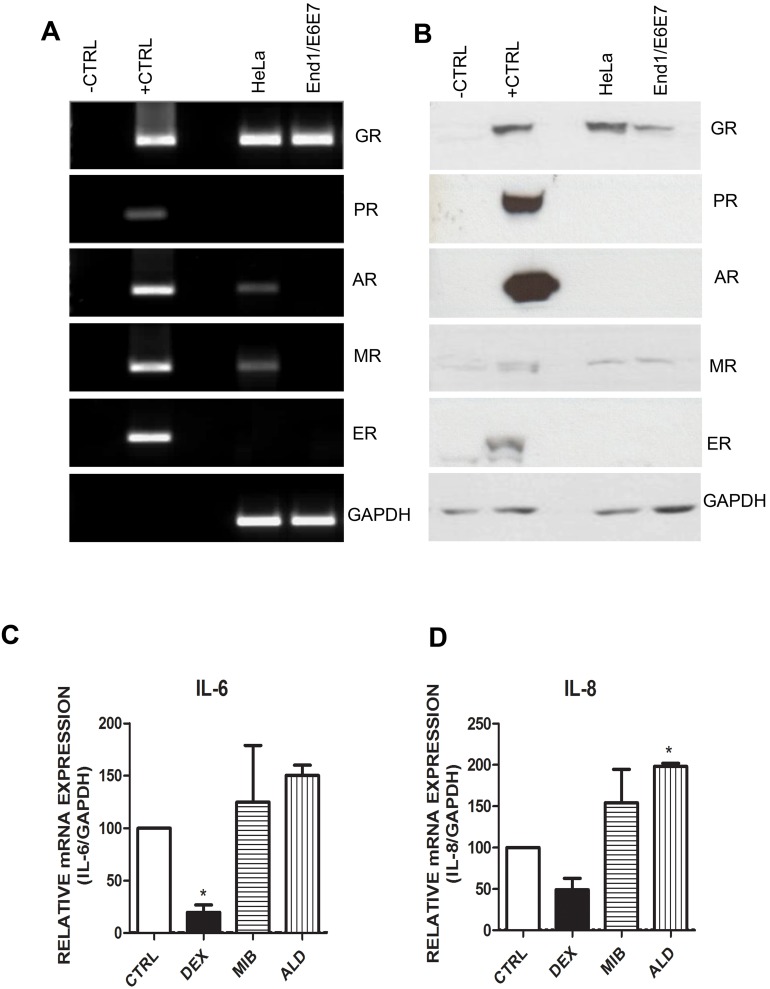
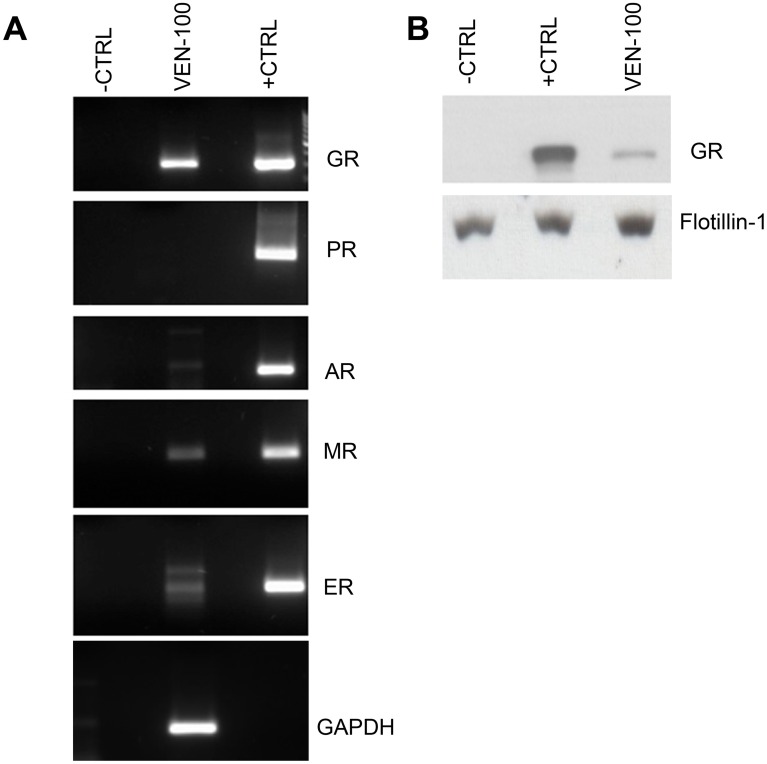
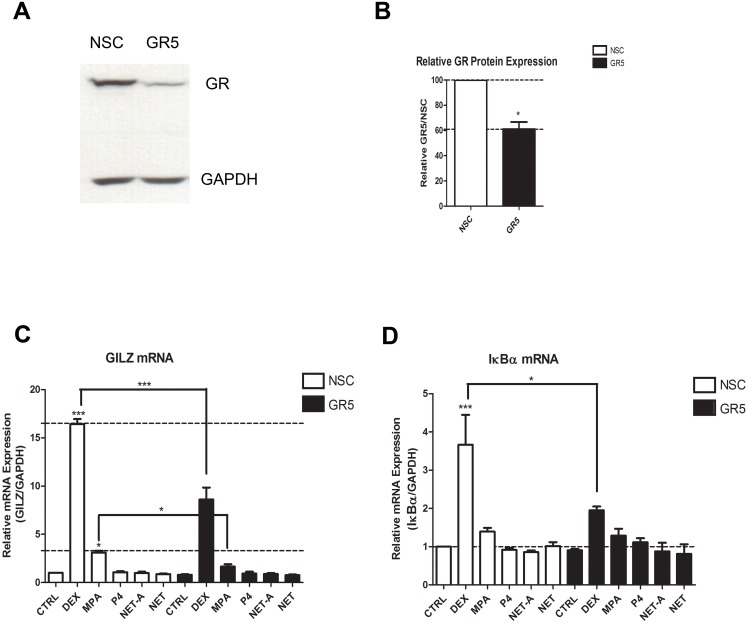
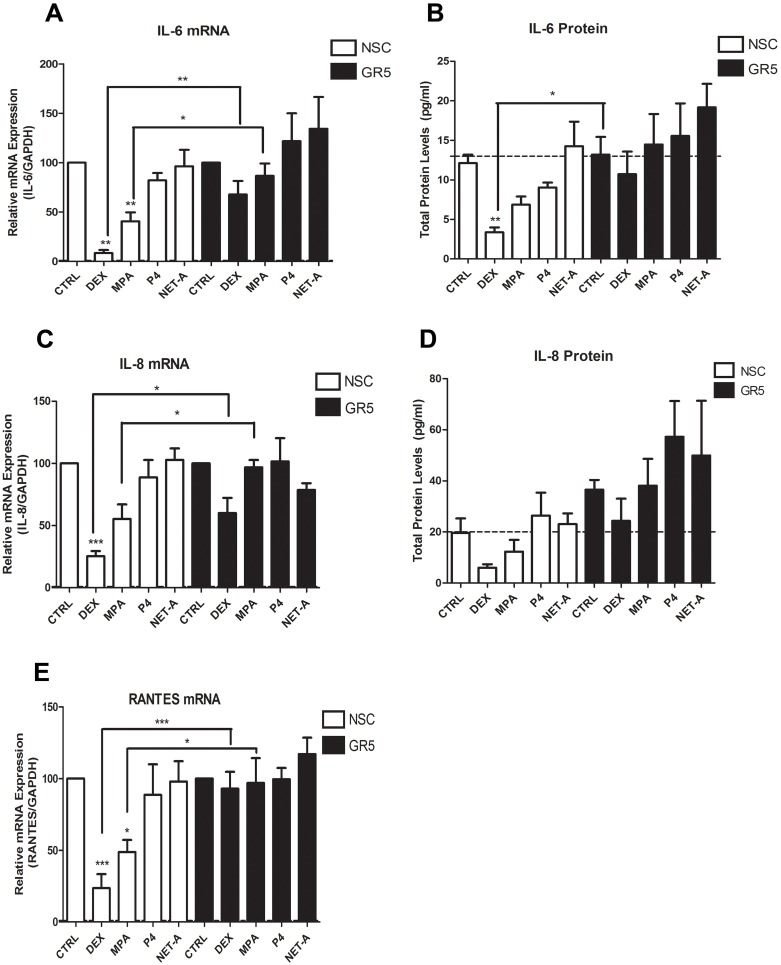
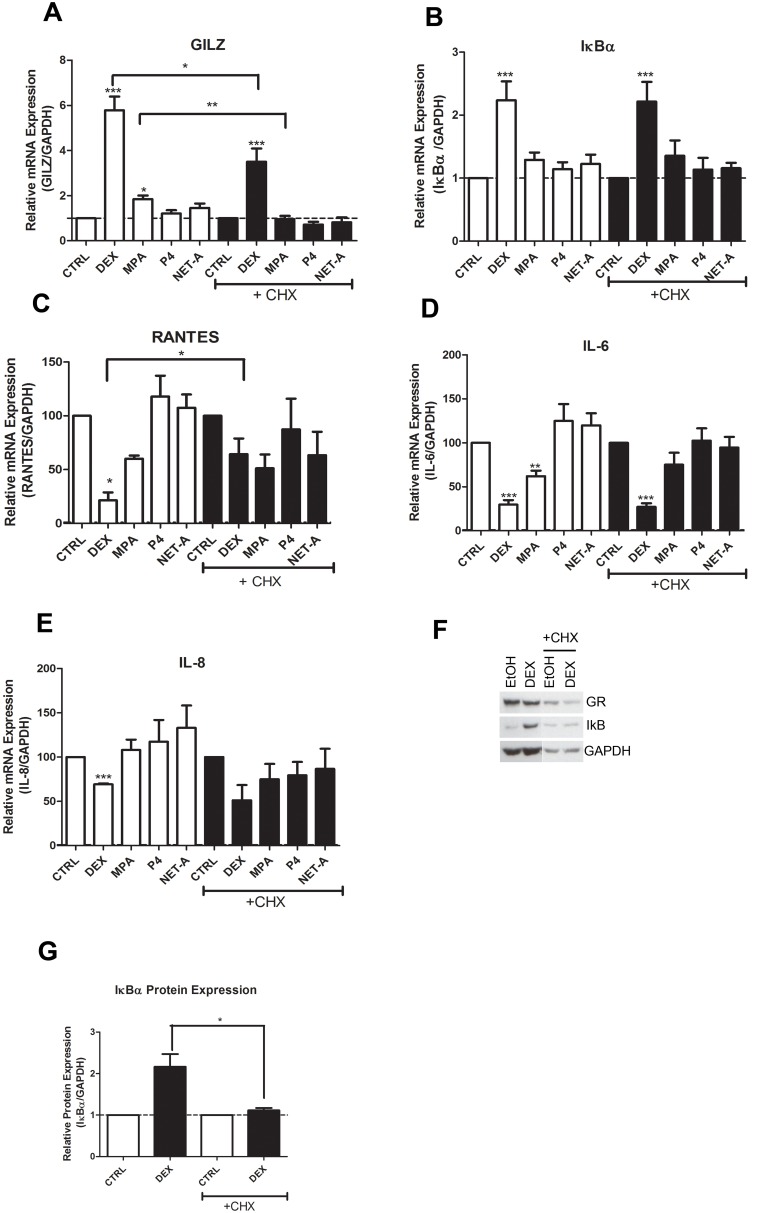
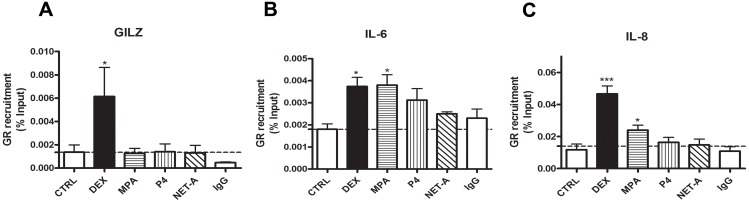
Clinical studies suggest that the injectable contraceptive medroxyprogesterone acetate (MPA) increases susceptibility to infections such as HIV-1, unlike the injectable contraceptive norethisterone enanthate (NET-EN). We investigated the differential effects, molecular mechanism of action and steroid receptor involvement in gene expression by MPA as compared to NET and progesterone (P4) in the End1/E6E7 cell line model for the endocervical epithelium, a key point of entry for pathogens in the female genital mucosa. MPA, unlike NET-acetate (NET-A) and P4, increases mRNA expression of the anti-inflammatory GILZ and IκBα genes. Similarly, MPA unlike NET-A, decreases mRNA expression of the pro-inflammatory IL-6, IL-8 and RANTES genes, and IL-6 and IL-8 protein levels. The predominant steroid receptor expressed in the End1/E6E7 and primary endocervical epithelial cells is the glucocorticoid receptor (GR), and GR knockdown experiments show that the anti-inflammatory effects of MPA are mediated by the GR. Chromatin-immunoprecipitation results suggest that MPA, unlike NET-A and P4, represses pro-inflammatory cytokine gene expression in cervical epithelial cells via a mechanism involving recruitment of the GR to cytokine gene promoters, like the GR agonist dexamethasone. This is at least in part consistent with direct effects on transcription, without a requirement for new protein synthesis. Dose response analysis shows that MPA has a potency of ∼ 24 nM for transactivation of the anti-inflammatory GILZ gene and ∼ 4-20 nM for repression of the pro-inflammatory genes, suggesting that these effects are likely to be relevant at injectable contraceptive doses of MPA. These findings suggest that in the context of the genital mucosa, these GR-mediated glucocorticoid-like effects of MPA in cervical epithelial cells are likely to play a critical role in discriminating between the effects on inflammation caused by different progestins and P4 and hence susceptibility to genital infections, given the predominant expression of the GR in primary endocervical epithelial cells.
Conflict of interest statement
Figures
Similar articles
-
The progestin-only contraceptive medroxyprogesterone acetate, but not norethisterone acetate, enhances HIV-1 Vpr-mediated apoptosis in human CD4+ T cells through the glucocorticoid receptor.PLoS One. 2013 May 3;8(5):e62895. doi: 10.1371/journal.pone.0062895. Print 2013. PLoS One. 2013. PMID: 23658782 Free PMC article.
-
Differential regulation of endogenous pro-inflammatory cytokine genes by medroxyprogesterone acetate and norethisterone acetate in cell lines of the female genital tract.Contraception. 2011 Oct;84(4):423-35. doi: 10.1016/j.contraception.2011.06.006. Epub 2011 Aug 4. Contraception. 2011. PMID: 21920200
-
Differential glucocorticoid receptor-mediated effects on immunomodulatory gene expression by progestin contraceptives: implications for HIV-1 pathogenesis.Am J Reprod Immunol. 2014 Jun;71(6):505-12. doi: 10.1111/aji.12214. Epub 2014 Feb 18. Am J Reprod Immunol. 2014. PMID: 24547700 Review.
-
Medroxyprogesterone acetate differentially regulates interleukin (IL)-12 and IL-10 in a human ectocervical epithelial cell line in a glucocorticoid receptor (GR)-dependent manner.J Biol Chem. 2014 Nov 7;289(45):31136-49. doi: 10.1074/jbc.M114.587311. Epub 2014 Sep 8. J Biol Chem. 2014. PMID: 25202013 Free PMC article.
-
Pharmacodynamic effects of once-a-month combined injectable contraceptives.Contraception. 1994 Apr;49(4):361-85. doi: 10.1016/0010-7824(94)90033-7. Contraception. 1994. PMID: 8013220 Review.
Cited by
-
Use of contraceptive depot medroxyprogesterone acetate is associated with impaired cervicovaginal mucosal integrity.J Clin Invest. 2018 Oct 1;128(10):4622-4638. doi: 10.1172/JCI120583. Epub 2018 Sep 17. J Clin Invest. 2018. PMID: 30222141 Free PMC article. Clinical Trial.
-
Differential metabolism of clinically-relevant progestogens in cell lines and tissue: Implications for biological mechanisms.J Steroid Biochem Mol Biol. 2019 May;189:145-153. doi: 10.1016/j.jsbmb.2019.02.010. Epub 2019 Feb 26. J Steroid Biochem Mol Biol. 2019. PMID: 30822501 Free PMC article.
-
Changes in Vaginal Microbiota and Immune Mediators in HIV-1-Seronegative Kenyan Women Initiating Depot Medroxyprogesterone Acetate.J Acquir Immune Defic Syndr. 2016 Apr 1;71(4):359-66. doi: 10.1097/QAI.0000000000000866. J Acquir Immune Defic Syndr. 2016. PMID: 26914908 Free PMC article.
-
Studying the effects of reproductive hormones and bacterial vaginosis on the glycome of lavage samples from the cervicovaginal cavity.PLoS One. 2015 May 20;10(5):e0127021. doi: 10.1371/journal.pone.0127021. eCollection 2015. PLoS One. 2015. PMID: 25993513 Free PMC article.
-
Reciprocal Modulation of Antiretroviral Drug and Steroid Receptor Function In Vitro.Antimicrob Agents Chemother. 2019 Dec 20;64(1):e01890-19. doi: 10.1128/AAC.01890-19. Print 2019 Dec 20. Antimicrob Agents Chemother. 2019. PMID: 31658973 Free PMC article.
References
-
- Kaushic C, Ferreira VH, Kafka JK, Nazli A (2010) HIV infection in the female genital tract: discrete influence of the local mucosal microenvironment. Am J Reprod Immunol 63: 566–575. - PubMed
-
- Kaushic C (2011) HIV-1 infection in the female reproductive tract: role of interactions between HIV-1 and genital epithelial cells. Am J Reprod Immunol 65: 253–260. - PubMed
-
- Wira CR, Fahey JV, Sentman CL, Pioli PA, Shen L (2005) Innate and adaptive immunity in female genital tract: cellular responses and interactions. Immunol Rev 206: 306–335. - PubMed
-
- Barclay C, Brennand J, Kelly R, Calder A (1993) Interleukin-8 production by the human cervix. Am J Obs Gynecol 169: 625–632. - PubMed
-
- Fichorova RN, Anderson DJ (1999) Differential expression of immunobiological mediators by immortalized human cervical and vaginal epithelial cells. Biol Reprod 60: 508–514. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
