

Disulfide bond requirements for active Wnt ligands
- PMID: 24841207
- PMCID: PMC4140276
- DOI: 10.1074/jbc.M114.575027
Disulfide bond requirements for active Wnt ligands
Abstract
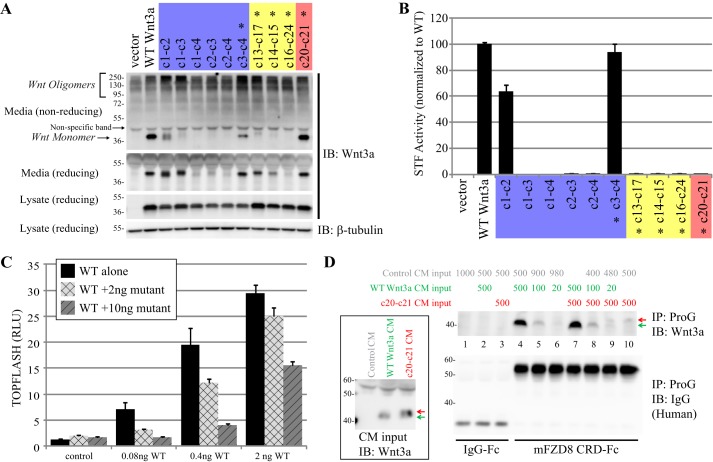
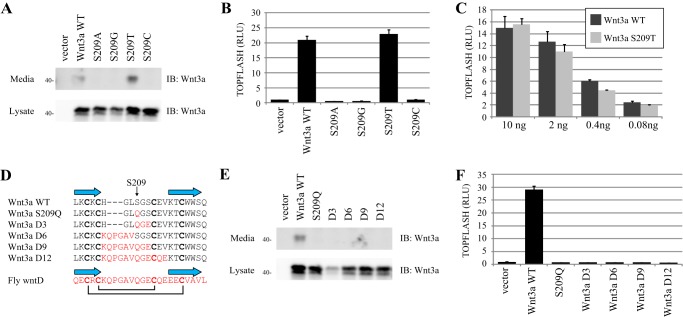
Secreted Wnt lipoproteins are cysteine-rich and lipid-modified morphogens that bind to the Frizzled (FZD) receptor and LDL receptor-related protein 6 (LRP6). Wnt engages FZD through protruding thumb and index finger domains, which are each assembled from paired β strands secured by disulfide bonds and grasp two sides of the FZD ectodomain. The importance of Wnt disulfide bonds has been assumed but uncharacterized. We systematically analyzed cysteines and associated disulfide bonds in the prototypic Wnt3a. Our data show that mutation of any individual cysteine of Wnt3a results in covalent Wnt oligomers through ectopic intermolecular disulfide bond formation and diminishes/abolishes Wnt signaling. Although individual cysteine mutations in the amino part of the saposin-like domain and in the base of the index finger are better tolerated and permit residual Wnt3a secretion/activity, those in the amino terminus, the thumb, and at the tip of the index finger are incompatible with secretion and/or activity. A few select double cysteine mutants based on the disulfide bond pattern restore Wnt secretion/activity. Further, a double cysteine mutation at the index finger tip results in a Wnt3a with normal secretion but minimal FZD binding and dominant negative properties. Our results experimentally validate predictions from the Wnt crystal structure and highlight critical but different roles of the saposin-like and cytokine-like domains, including the thumb and the index finger in Wnt folding/secretion and FZD binding. Finally, we modified existing expression vectors for 19 epitope-tagged human WNT proteins by removal of a tag-supplied ectopic cysteine, thereby generating tagged WNT ligands active in canonical and non-canonical signaling.
Keywords: Disulfide; Protein Domain; Wnt Biogenesis; Wnt Pathway; Wnt Signaling; Wnt3a; beta-Catenin.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures




Similar articles
-
The role of Wnt palmitoleylated loop conserved disulfide bonds in Wnt-frizzled complex structural dynamics: Insights from molecular dynamics simulations.Comput Biol Med. 2023 Dec;167:107703. doi: 10.1016/j.compbiomed.2023.107703. Epub 2023 Nov 11. Comput Biol Med. 2023. PMID: 37979393
-
Secreted Frizzled-related protein potentiation versus inhibition of Wnt3a/β-catenin signaling.Cell Signal. 2014 Jan;26(1):94-101. doi: 10.1016/j.cellsig.2013.09.016. Epub 2013 Sep 28. Cell Signal. 2014. PMID: 24080158 Free PMC article.
-
β-Catenin-dependent pathway activation by both promiscuous "canonical" WNT3a-, and specific "noncanonical" WNT4- and WNT5a-FZD receptor combinations with strong differences in LRP5 and LRP6 dependency.Cell Signal. 2014 Feb;26(2):260-7. doi: 10.1016/j.cellsig.2013.11.021. Epub 2013 Nov 21. Cell Signal. 2014. PMID: 24269653
-
Fatty acid recognition in the Frizzled receptor family.J Biol Chem. 2019 Jan 11;294(2):726-736. doi: 10.1074/jbc.REV118.005205. Epub 2018 Dec 10. J Biol Chem. 2019. PMID: 30530496 Free PMC article. Review.
-
Functionally annotating cysteine disulfides and metal binding sites in the plant kingdom using AlphaFold2 predicted structures.Free Radic Biol Med. 2023 Jan;194:220-229. doi: 10.1016/j.freeradbiomed.2022.12.001. Epub 2022 Dec 7. Free Radic Biol Med. 2023. PMID: 36493985 Review.
Cited by
-
Wnt2b attenuates HSCs activation and liver fibrosis through negative regulating TLR4 signaling.Sci Rep. 2017 Jun 21;7(1):3952. doi: 10.1038/s41598-017-04374-5. Sci Rep. 2017. PMID: 28638086 Free PMC article.
-
Noncanonical WNT-5A signaling impairs endogenous lung repair in COPD.J Exp Med. 2017 Jan;214(1):143-163. doi: 10.1084/jem.20160675. Epub 2016 Dec 15. J Exp Med. 2017. PMID: 27979969 Free PMC article.
-
WNT5a export onto extracellular vesicles studied at single-molecule and single-vesicle resolution.FEBS J. 2025 Sep;292(17):4631-4649. doi: 10.1111/febs.70074. Epub 2025 Apr 1. FEBS J. 2025. PMID: 40165582 Free PMC article.
-
Wnt-5A/B Signaling in Hematopoiesis throughout Life.Cells. 2020 Jul 29;9(8):1801. doi: 10.3390/cells9081801. Cells. 2020. PMID: 32751131 Free PMC article. Review.
-
Essential cell-extrinsic requirement for PDIA6 in lymphoid and myeloid development.J Exp Med. 2020 Apr 6;217(4):e20190006. doi: 10.1084/jem.20190006. J Exp Med. 2020. PMID: 31985756 Free PMC article.
References
-
- Clevers H., Nusse R. (2012) Wnt/β-catenin signaling and disease. Cell 149, 1192–1205 - PubMed
-
- Anastas J. N., Moon R. T. (2013) WNT signalling pathways as therapeutic targets in cancer. Nat. Rev. Cancer 13, 11–26 - PubMed
-
- Coudreuse D., Korswagen H. C. (2007) The making of Wnt: new insights into Wnt maturation, sorting and secretion. Development 134, 3–12 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
