

Regulated proteolysis of NOTCH2 and NOTCH3 receptors by ADAM10 and presenilins
- PMID: 24842903
- PMCID: PMC4135574
- DOI: 10.1128/MCB.00206-14
Regulated proteolysis of NOTCH2 and NOTCH3 receptors by ADAM10 and presenilins
Abstract
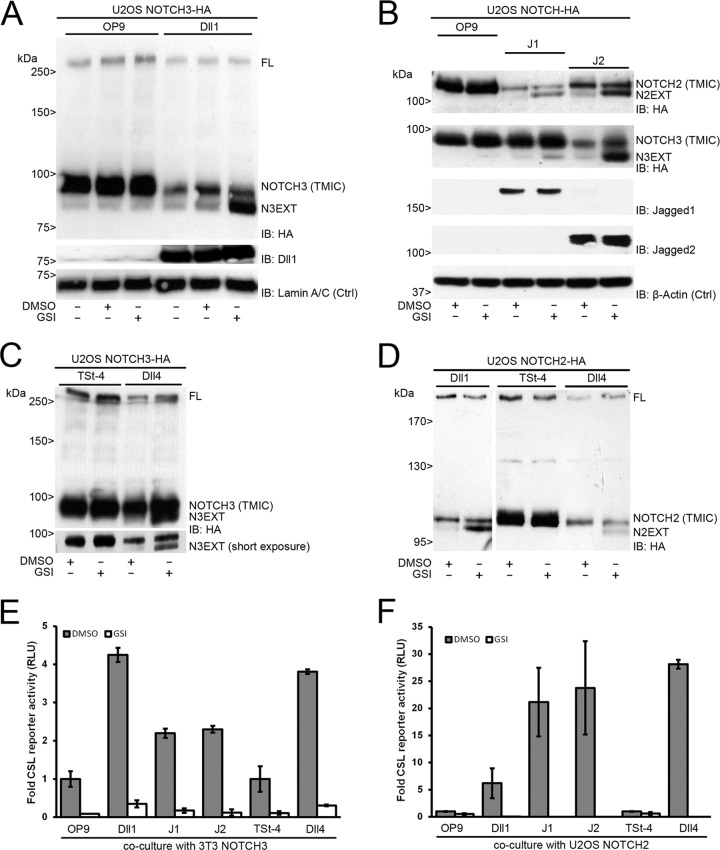
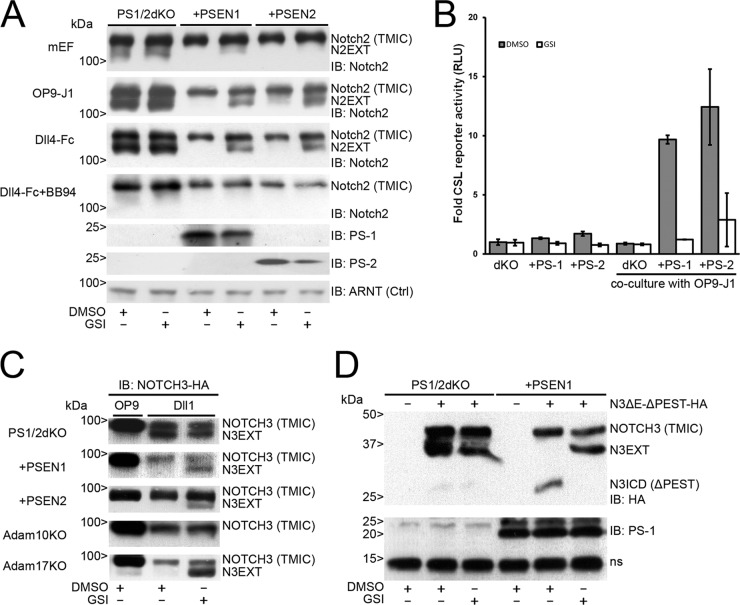
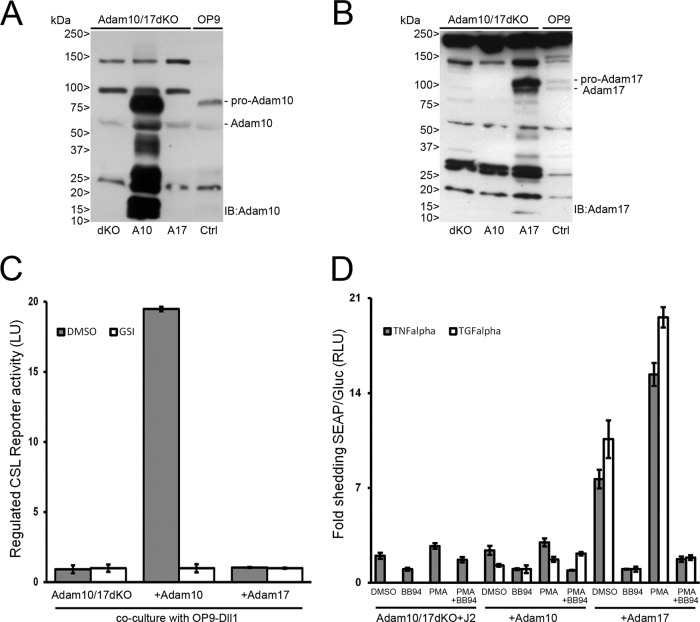
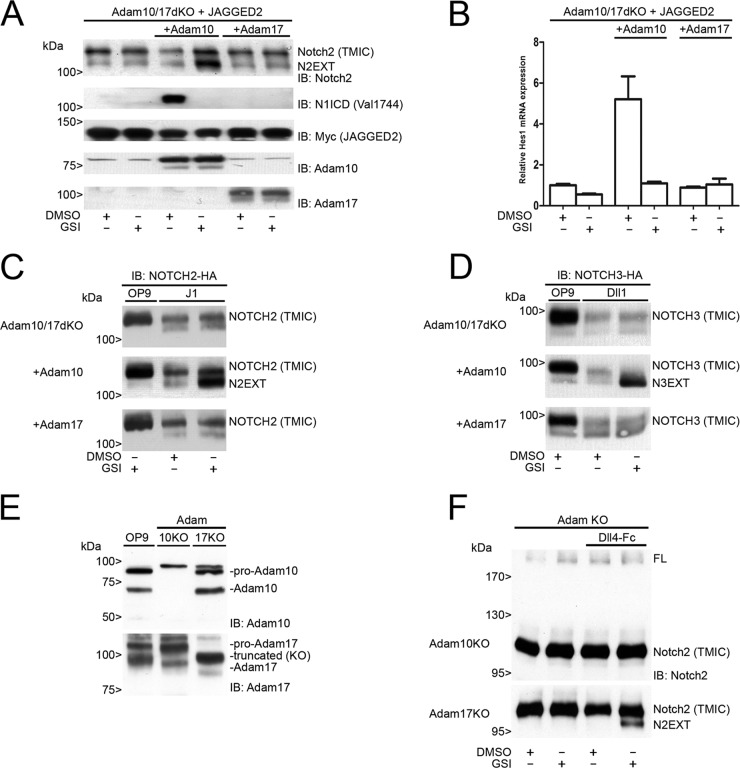
In mammals, there are four NOTCH receptors and five Delta-Jagged-type ligands regulating many aspects of embryonic development and adult tissue homeostasis. NOTCH proteins are type I transmembrane receptors that interact with ligands on adjacent cells and are activated by regulated intramembrane proteolysis (RIP). The activation mechanism of NOTCH1 receptors upon ligand binding is well understood and requires cleavage by ADAM10 metalloproteases prior to intramembranous cleavage by γ-secretase. How the other human NOTCH receptor homologues are activated upon ligand binding is not known. Here, we dissect the proteolytic activation mechanism of the NOTCH2 and NOTCH3 receptors. We show that NOTCH2 and NOTCH3 signaling can be triggered by both Delta-Jagged-type ligands and requires ADAM10 and presenilin-1 or -2. Importantly, we did not find any role for the highly related ADAM17/TACE (tumor necrosis factor alpha-converting enzyme) protease in ligand-induced NOTCH2 or NOTCH3 signaling. These results demonstrate that canonical ligand-induced proteolysis of the NOTCH1, -2, and -3 receptors strictly depends on consecutive cleavage of these receptors by ADAM10 and the presenilin-containing γ-secretase complex, leading to transcriptional activation.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Human NOTCH2 Is Resistant to Ligand-independent Activation by Metalloprotease Adam17.J Biol Chem. 2015 Jun 5;290(23):14705-16. doi: 10.1074/jbc.M115.643676. Epub 2015 Apr 27. J Biol Chem. 2015. PMID: 25918160 Free PMC article.
-
The Notch ligands, Jagged and Delta, are sequentially processed by alpha-secretase and presenilin/gamma-secretase and release signaling fragments.J Biol Chem. 2003 Sep 5;278(36):34427-37. doi: 10.1074/jbc.M302659200. Epub 2003 Jun 25. J Biol Chem. 2003. PMID: 12826675
-
ADAM10, the rate-limiting protease of regulated intramembrane proteolysis of Notch and other proteins, is processed by ADAMS-9, ADAMS-15, and the gamma-secretase.J Biol Chem. 2009 Apr 24;284(17):11738-47. doi: 10.1074/jbc.M805894200. Epub 2009 Feb 11. J Biol Chem. 2009. PMID: 19213735 Free PMC article.
-
Implication of APP secretases in notch signaling.J Mol Neurosci. 2001 Oct;17(2):171-81. doi: 10.1385/JMN:17:2:171. J Mol Neurosci. 2001. PMID: 11816790 Review.
-
gamma-Secretase: a multifaceted regulator of angiogenesis.J Cell Mol Med. 2008 Jun;12(3):781-95. doi: 10.1111/j.1582-4934.2008.00274.x. Epub 2008 Feb 8. J Cell Mol Med. 2008. PMID: 18266961 Free PMC article. Review.
Cited by
-
Notch signaling promotes osteoclast maturation and resorptive activity.J Cell Biochem. 2015 Nov;116(11):2598-609. doi: 10.1002/jcb.25205. J Cell Biochem. 2015. PMID: 25914241 Free PMC article.
-
Secretases Related to Amyloid Precursor Protein Processing.Membranes (Basel). 2021 Dec 15;11(12):983. doi: 10.3390/membranes11120983. Membranes (Basel). 2021. PMID: 34940484 Free PMC article. Review.
-
The Abelson kinase and the Nedd4 family E3 ligases co-regulate Notch trafficking to limit signaling.J Cell Biol. 2025 Jun 2;224(6):e202407066. doi: 10.1083/jcb.202407066. Epub 2025 Apr 4. J Cell Biol. 2025. PMID: 40183942
-
Genetic and polygenic risk score analysis for Alzheimer's disease in the Chinese population.Alzheimers Dement (Amst). 2020 Aug 5;12(1):e12074. doi: 10.1002/dad2.12074. eCollection 2020. Alzheimers Dement (Amst). 2020. PMID: 32775599 Free PMC article.
-
Targeting Notch to overcome radiation resistance.Oncotarget. 2016 Feb 16;7(7):7610-28. doi: 10.18632/oncotarget.6714. Oncotarget. 2016. PMID: 26713603 Free PMC article. Review.
References
-
- Sanchez-Irizarry C, Carpenter AC, Weng AP, Pear WS, Aster JC, Blacklow SC. 2004. Notch subunit heterodimerization and prevention of ligand-independent proteolytic activation depend, respectively, on a novel domain and the LNR repeats. Mol. Cell. Biol. 24:9265–9273. 10.1128/MCB.24.21.9265-9273.2004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous