

Recombinogenic telomeres in diploid Sorex granarius (Soricidae, Eulipotyphla) fibroblast cells
- PMID: 24842907
- PMCID: PMC4135569
- DOI: 10.1128/MCB.01697-13
Recombinogenic telomeres in diploid Sorex granarius (Soricidae, Eulipotyphla) fibroblast cells
Abstract
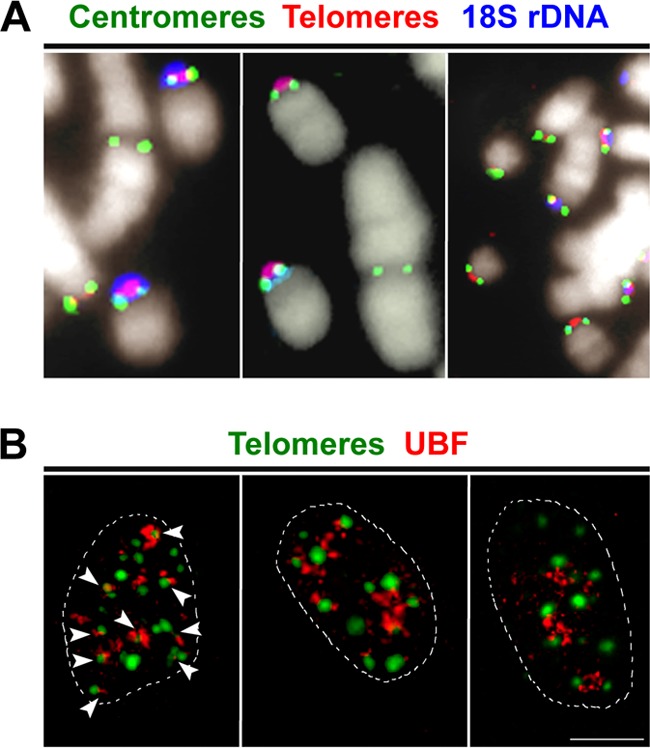
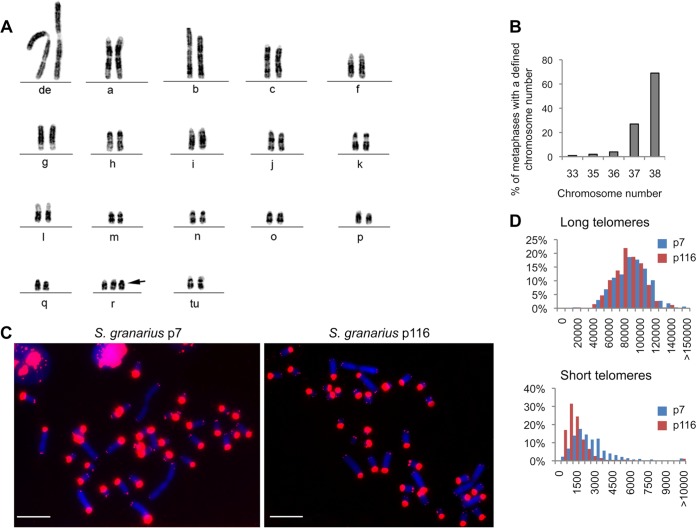
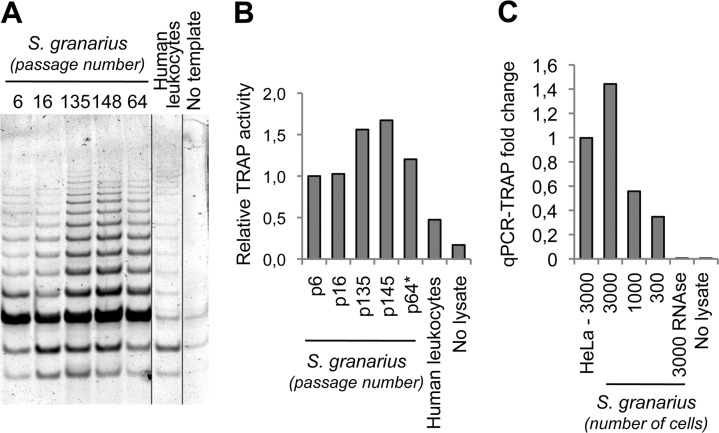
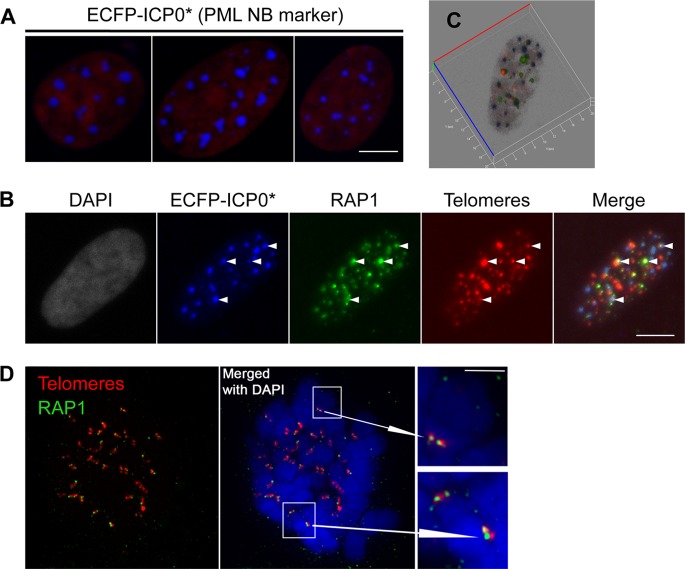
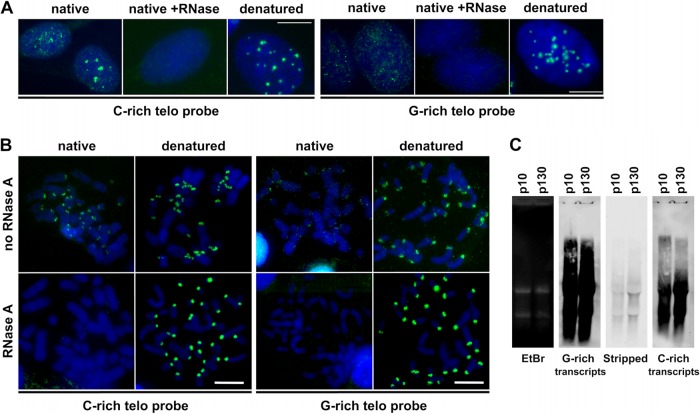
The telomere structure in the Iberian shrew Sorex granarius is characterized by unique, striking features, with short arms of acrocentric chromosomes carrying extremely long telomeres (up to 300 kb) with interspersed ribosomal DNA (rDNA) repeat blocks. In this work, we investigated the telomere physiology of S. granarius fibroblast cells and found that telomere repeats are transcribed on both strands and that there is no telomere-dependent senescence mechanism. Although telomerase activity is detectable throughout cell culture and appears to act on both short and long telomeres, we also discovered that signatures of a recombinogenic activity are omnipresent, including telomere-sister chromatid exchanges, formation of alternative lengthening of telomeres (ALT)-associated PML-like bodies, production of telomere circles, and a high frequency of telomeres carrying marks of a DNA damage response. Our results suggest that recombination participates in the maintenance of the very long telomeres in normal S. granarius fibroblasts. We discuss the possible interplay between the interspersed telomere and rDNA repeats in the stabilization of the very long telomeres in this organism.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
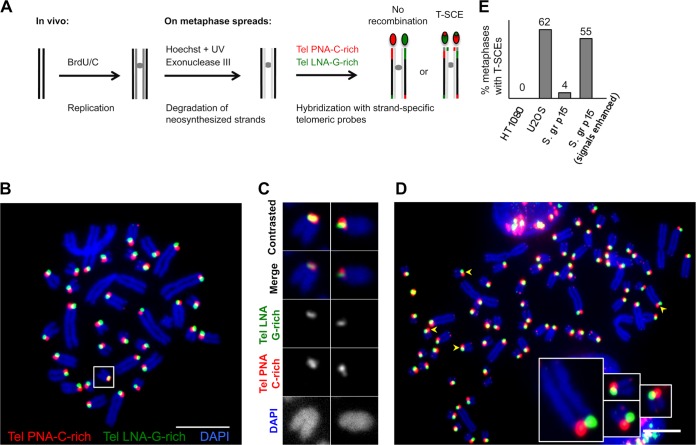
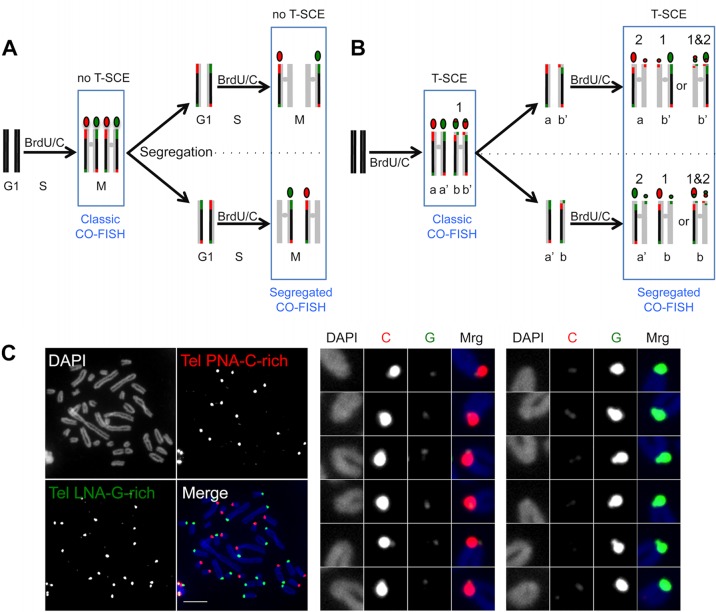
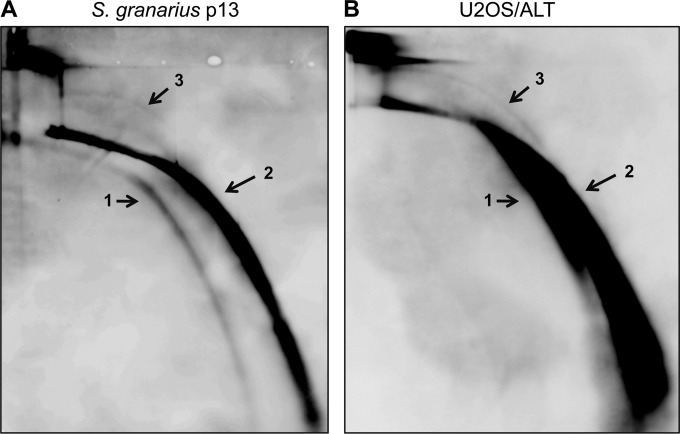
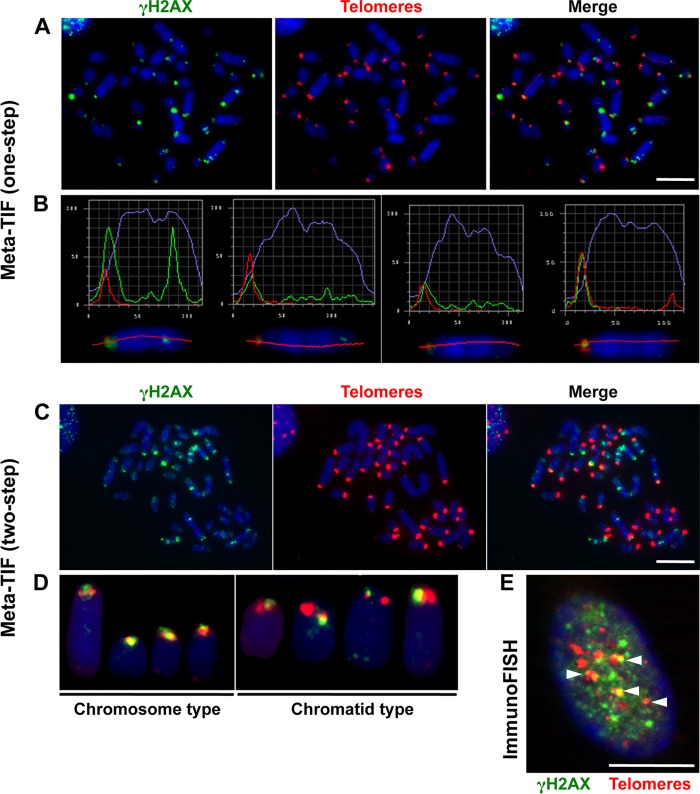
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources