

Ingestion of a moderate high-sucrose diet results in glucose intolerance with reduced liver glucokinase activity and impaired glucagon-like peptide-1 secretion
- PMID: 24843603
- PMCID: PMC4019243
- DOI: 10.1111/j.2040-1124.2012.00208.x
Ingestion of a moderate high-sucrose diet results in glucose intolerance with reduced liver glucokinase activity and impaired glucagon-like peptide-1 secretion
Abstract
Aims/Introduction: Excessive intake of sucrose can cause severe health issues, such as diabetes mellitus. In animal studies, consumption of a high-sucrose diet (SUC) has been shown to cause obesity, insulin resistance and glucose intolerance. However, several in vivo experiments have been carried out using diets with much higher sucrose contents (50-70% of the total calories) than are typically ingested by humans. In the present study, we examined the effects of a moderate SUC on glucose metabolism and the underlying mechanism.
Materials and methods: C57BL/6J mice received a SUC (38.5% sucrose), a high-starch diet (ST) or a control diet for 5 weeks. We assessed glucose tolerance, incretin secretion and liver glucose metabolism.
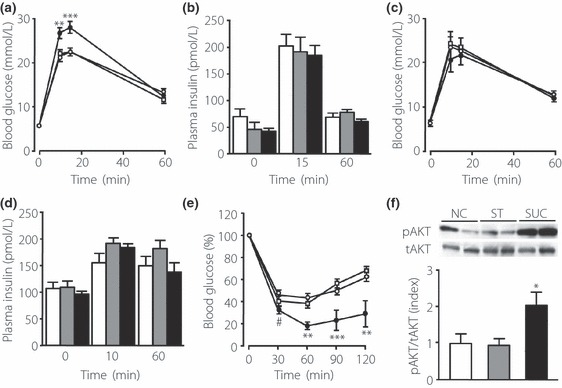
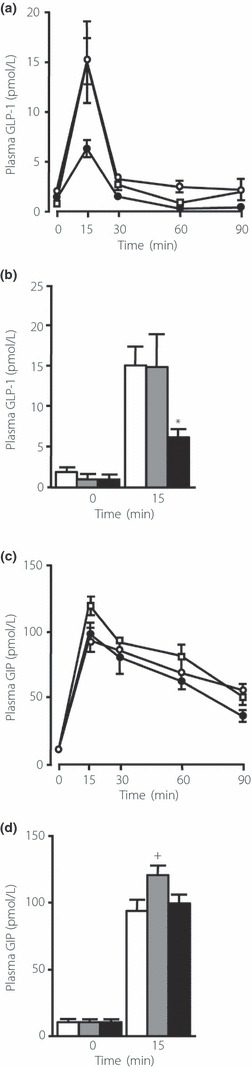
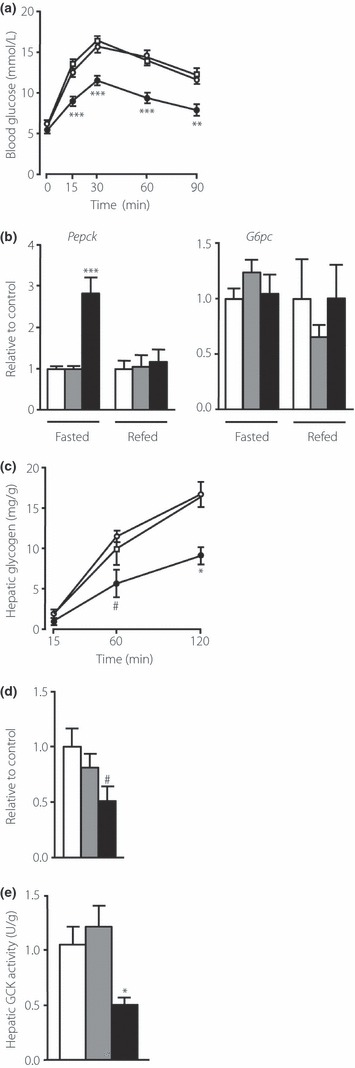
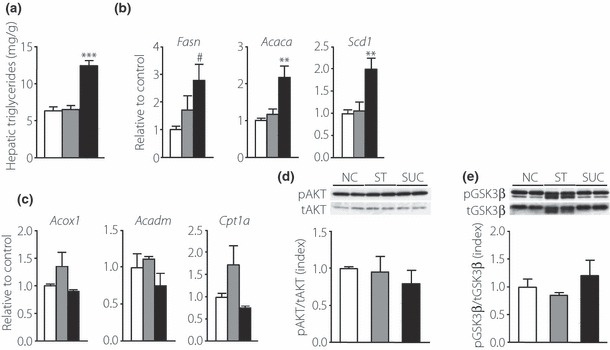
Results: An oral glucose tolerance test (OGTT) showed that plasma glucose levels in the early phase were significantly higher in SUC-fed mice than in ST-fed or control mice, with no change in plasma insulin levels at any stage. SUC-fed mice showed a significant improvement in insulin sensitivity. Glucagon-like peptide-1 (GLP-1) secretion 15 min after oral glucose administration was significantly lower in SUC-fed mice than in ST-fed or control mice. Hepatic glucokinase (GCK) activity was significantly reduced in SUC-fed mice. During the OGTT, the accumulation of glycogen in the liver was suppressed in SUC-fed mice in a time-dependent manner.
Conclusions: These results indicate that mice that consume a moderate SUC show glucose intolerance with a reduction in hepatic GCK activity and impairment in GLP-1 secretion. (J Diabetes Invest, doi: 10.1111/j.2040-1124.2012.00208.x, 2012).
Keywords: Glucagon‐like peptide‐1; Glucokinase; High‐sucrose diet.
Figures
References
-
- Laville M, Nazare JA. Diabetes, insulin resistance and sugars. Obes Rev 2009; 10(Suppl 1): 24–33 - PubMed
-
- Bray GA, Nielsen SJ, Popkin BM. Consumption of high‐fructose corn syrup in beverages may play a role in the epidemic of obesity. Am J Clin Nutr 2004; 79: 537–543 - PubMed
-
- Johnson RJ, Segal MS, Sautin Y, et al. Potential role of sugar (fructose) in the epidemic of hypertension, obesity and the metabolic syndrome, diabetes, kidney disease, and cardiovascular disease. Am J Clin Nutr 2007; 86: 899–906 - PubMed
-
- Pagliassotti MJ, Shahrokhi KA, Moscarello M. Involvement of liver and skeletal muscle in sucrose‐induced insulin resistance: dose‐response studies. Am J Physiol 1994; 266: R1637–R1644 - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
