

Structural and functional insights into the mode of action of a universally conserved Obg GTPase
- PMID: 24844575
- PMCID: PMC4028186
- DOI: 10.1371/journal.pbio.1001866
Structural and functional insights into the mode of action of a universally conserved Obg GTPase
Abstract
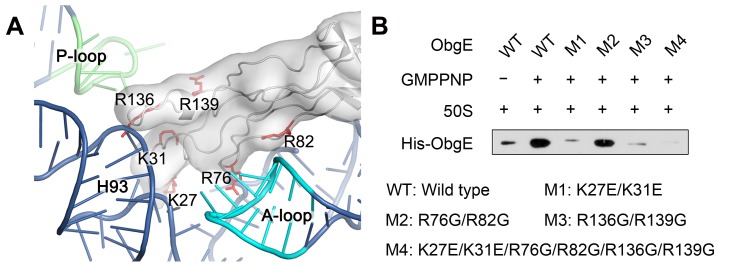
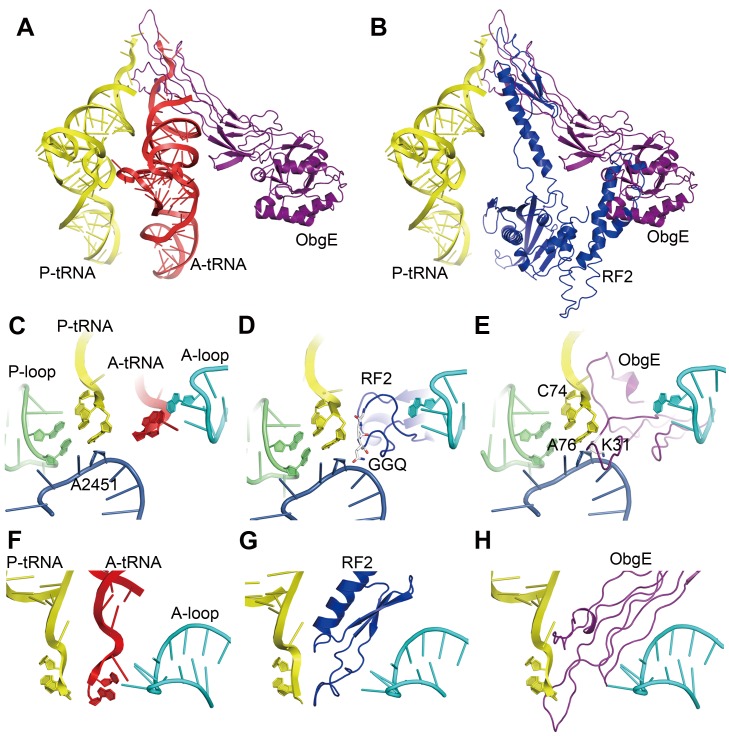
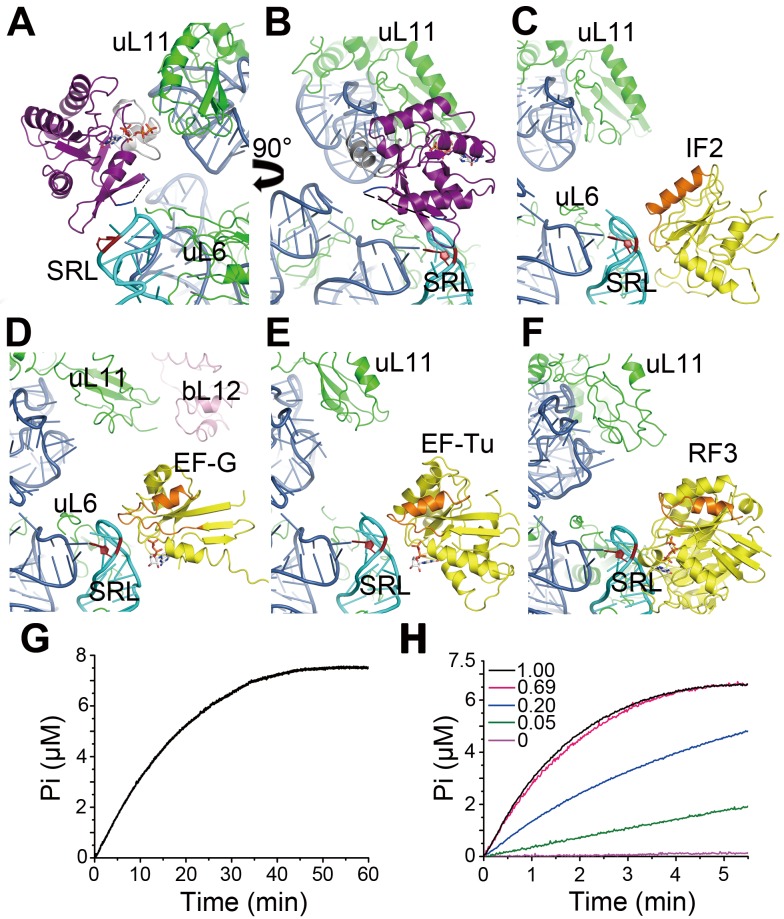
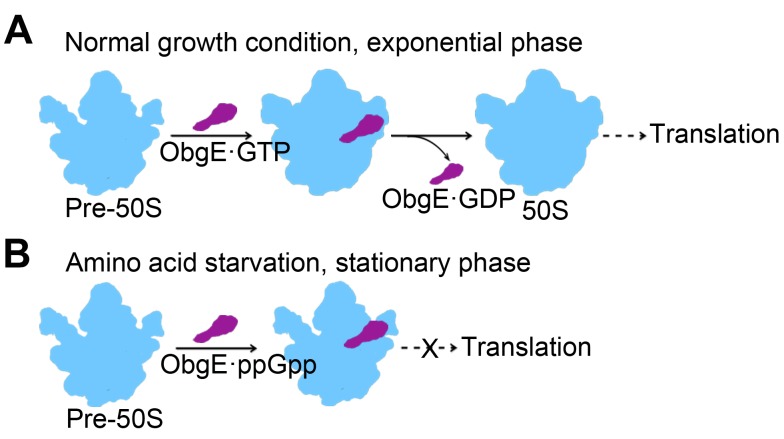
Obg proteins are a family of P-loop GTPases, conserved from bacteria to human. The Obg protein in Escherichia coli (ObgE) has been implicated in many diverse cellular functions, with proposed molecular roles in two global processes, ribosome assembly and stringent response. Here, using pre-steady state fast kinetics we demonstrate that ObgE is an anti-association factor, which prevents ribosomal subunit association and downstream steps in translation by binding to the 50S subunit. ObgE is a ribosome dependent GTPase; however, upon binding to guanosine tetraphosphate (ppGpp), the global regulator of stringent response, ObgE exhibits an enhanced interaction with the 50S subunit, resulting in increased equilibrium dissociation of the 70S ribosome into subunits. Furthermore, our cryo-electron microscopy (cryo-EM) structure of the 50S·ObgE·GMPPNP complex indicates that the evolutionarily conserved N-terminal domain (NTD) of ObgE is a tRNA structural mimic, with specific interactions with peptidyl-transferase center, displaying a marked resemblance to Class I release factors. These structural data might define ObgE as a specialized translation factor related to stress responses, and provide a framework towards future elucidation of functional interplay between ObgE and ribosome-associated (p)ppGpp regulators. Together with published data, our results suggest that ObgE might act as a checkpoint in final stages of the 50S subunit assembly under normal growth conditions. And more importantly, ObgE, as a (p)ppGpp effector, might also have a regulatory role in the production of the 50S subunit and its participation in translation under certain stressed conditions. Thus, our findings might have uncovered an under-recognized mechanism of translation control by environmental cues.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
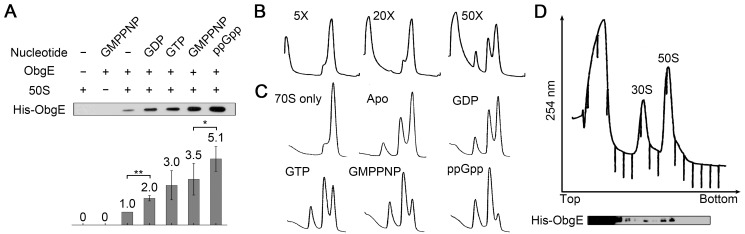
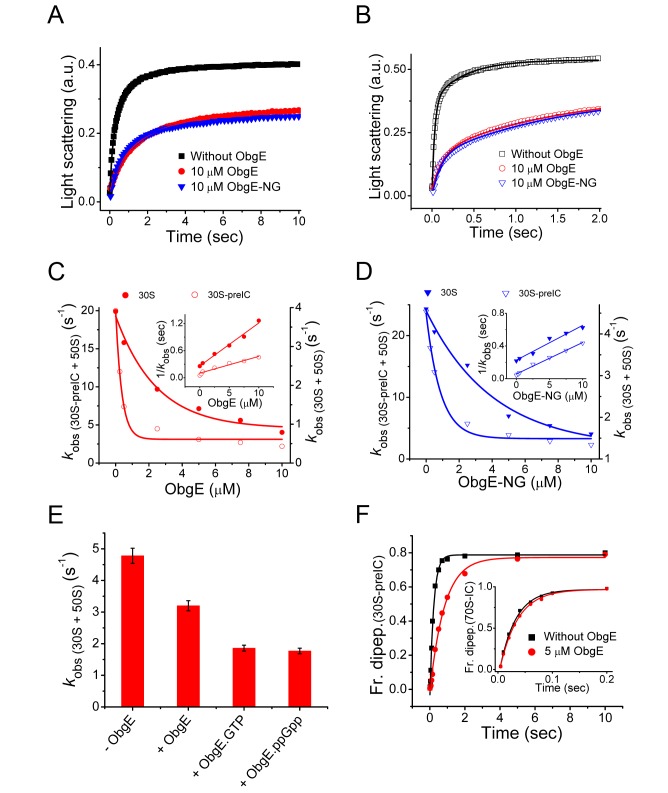
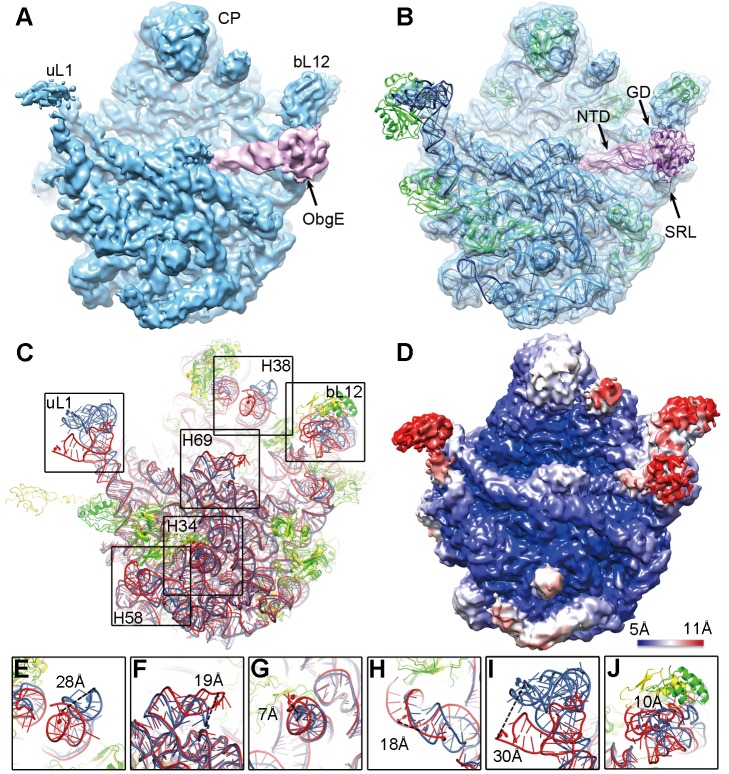
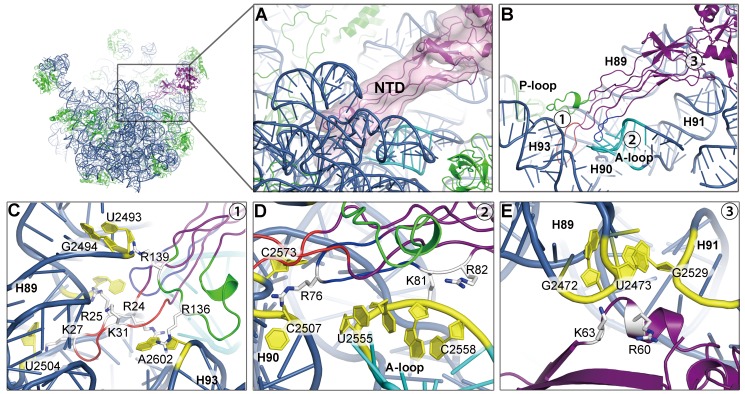
Comment in
-
Braking bad: stopping translation in hard times.PLoS Biol. 2014 May 20;12(5):e1001867. doi: 10.1371/journal.pbio.1001867. eCollection 2014 May. PLoS Biol. 2014. PMID: 24844695 Free PMC article. No abstract available.
References
-
- Wittinghofer A, Vetter IR (2011) Structure-function relationships of the G domain, a canonical switch motif. Annu Rev Biochem 80: 943–971. - PubMed
-
- Kint C, Verstraeten N, Hofkens J, Fauvart M, Michiels J (2013) Bacterial Obg proteins: GTPases at the nexus of protein and DNA synthesis. Crit Rev Microbiol. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
