

Mice deficient in transmembrane prostatic acid phosphatase display increased GABAergic transmission and neurological alterations
- PMID: 24846136
- PMCID: PMC4028278
- DOI: 10.1371/journal.pone.0097851
Mice deficient in transmembrane prostatic acid phosphatase display increased GABAergic transmission and neurological alterations
Abstract
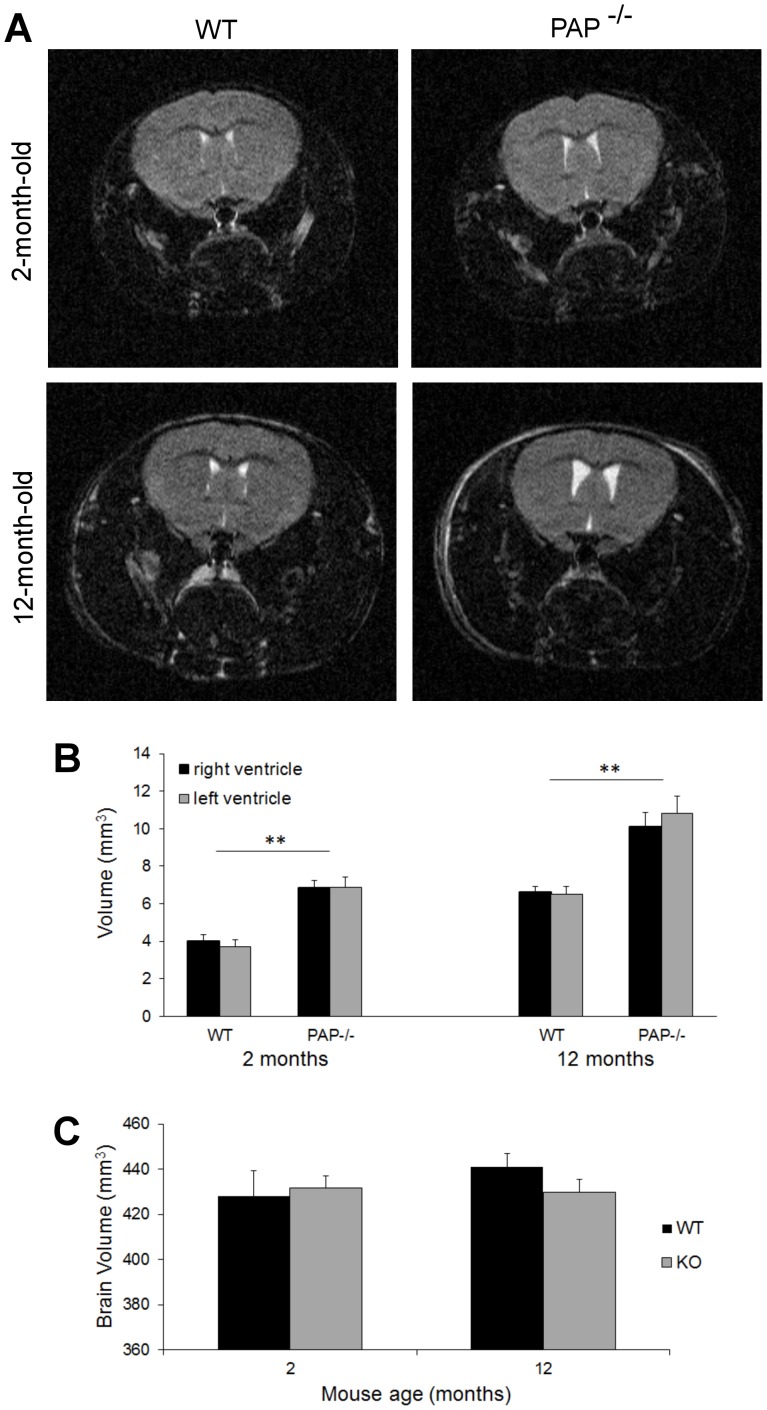
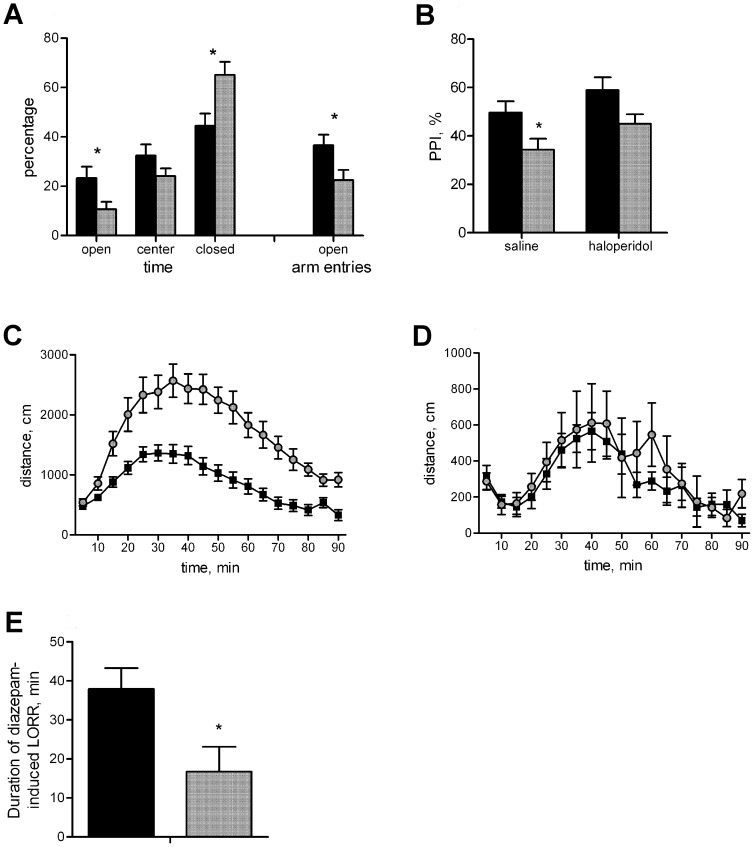
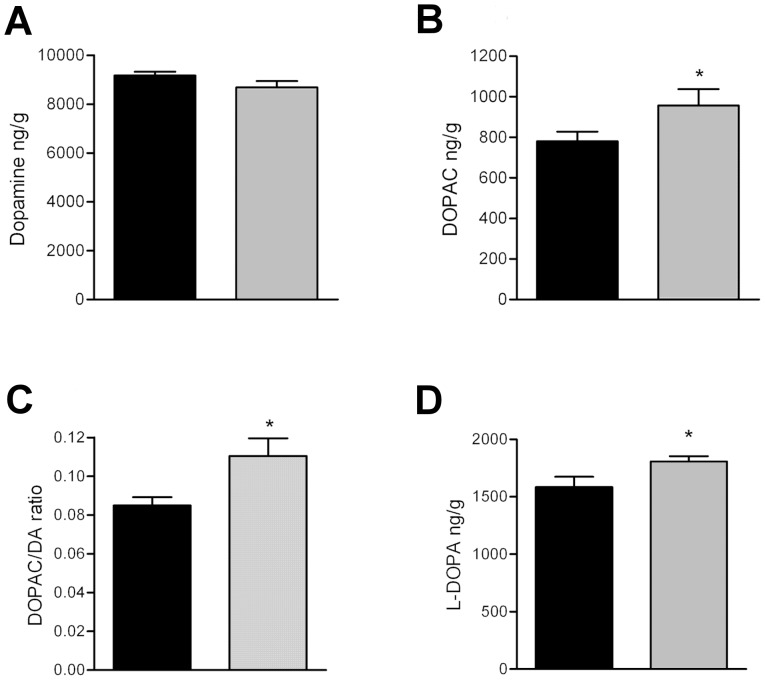
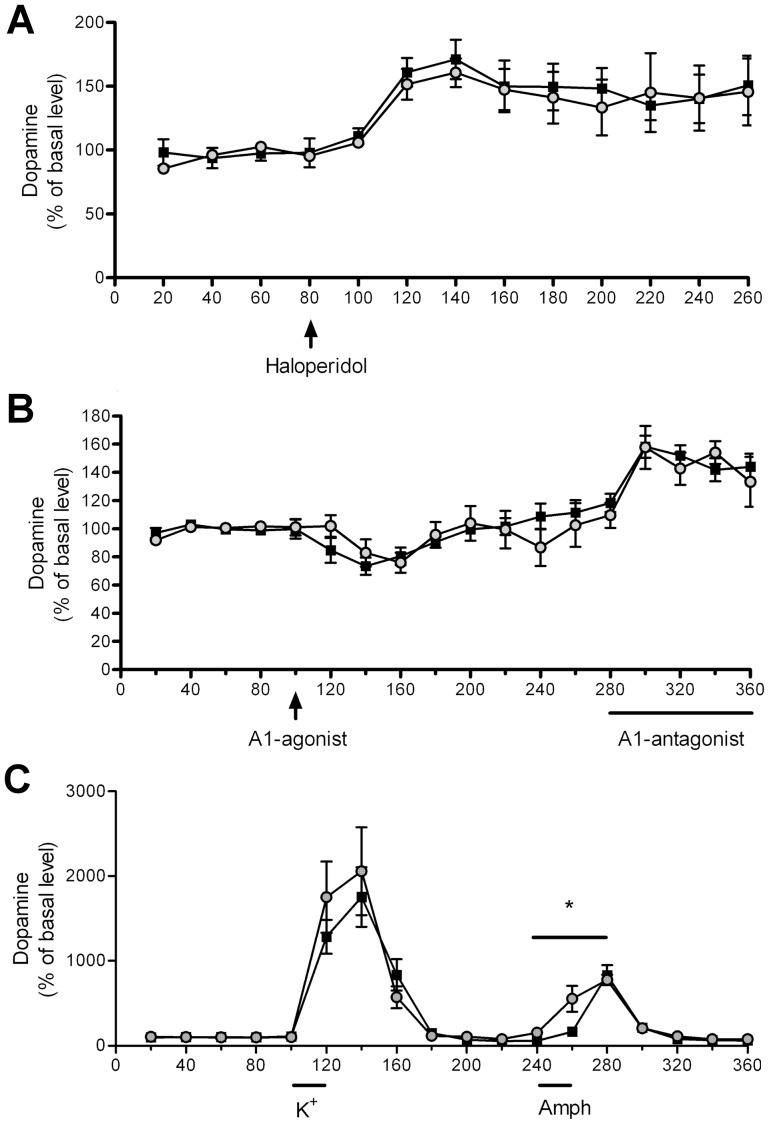
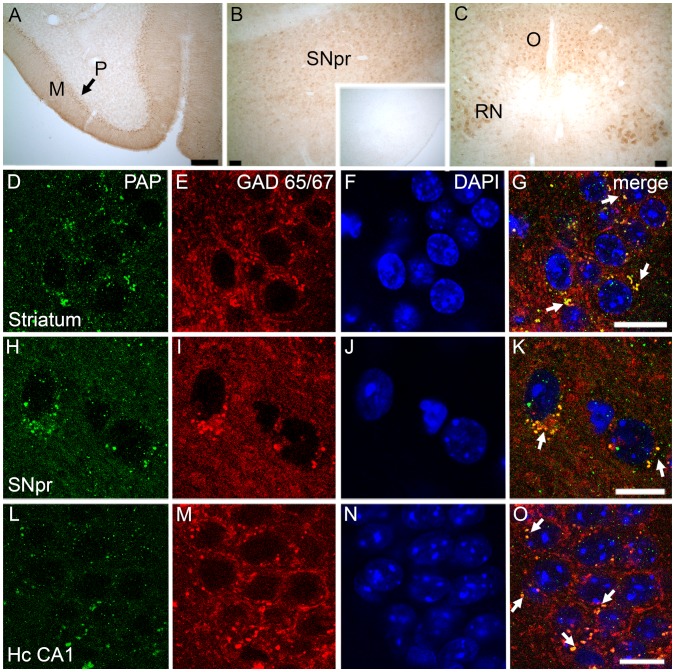
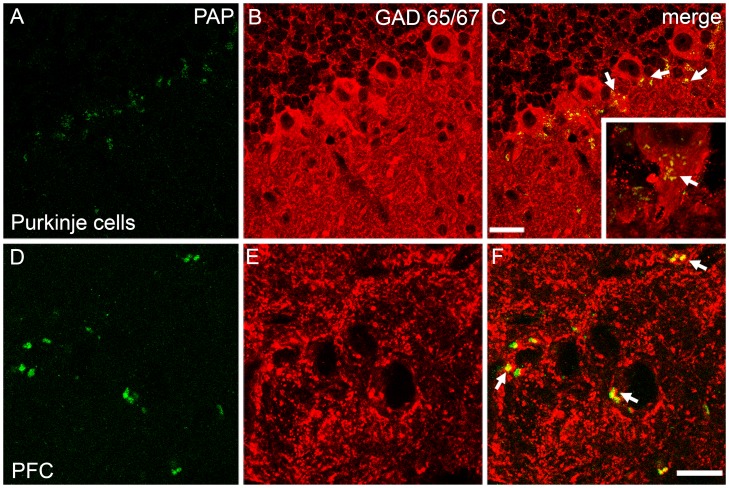
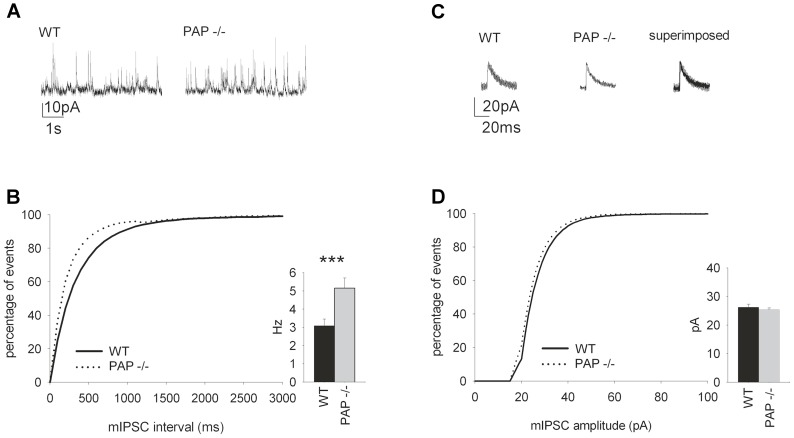
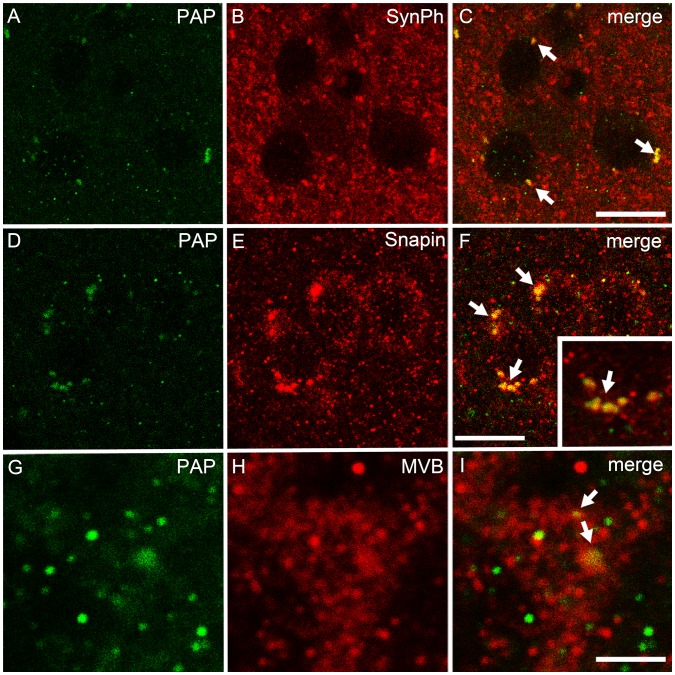
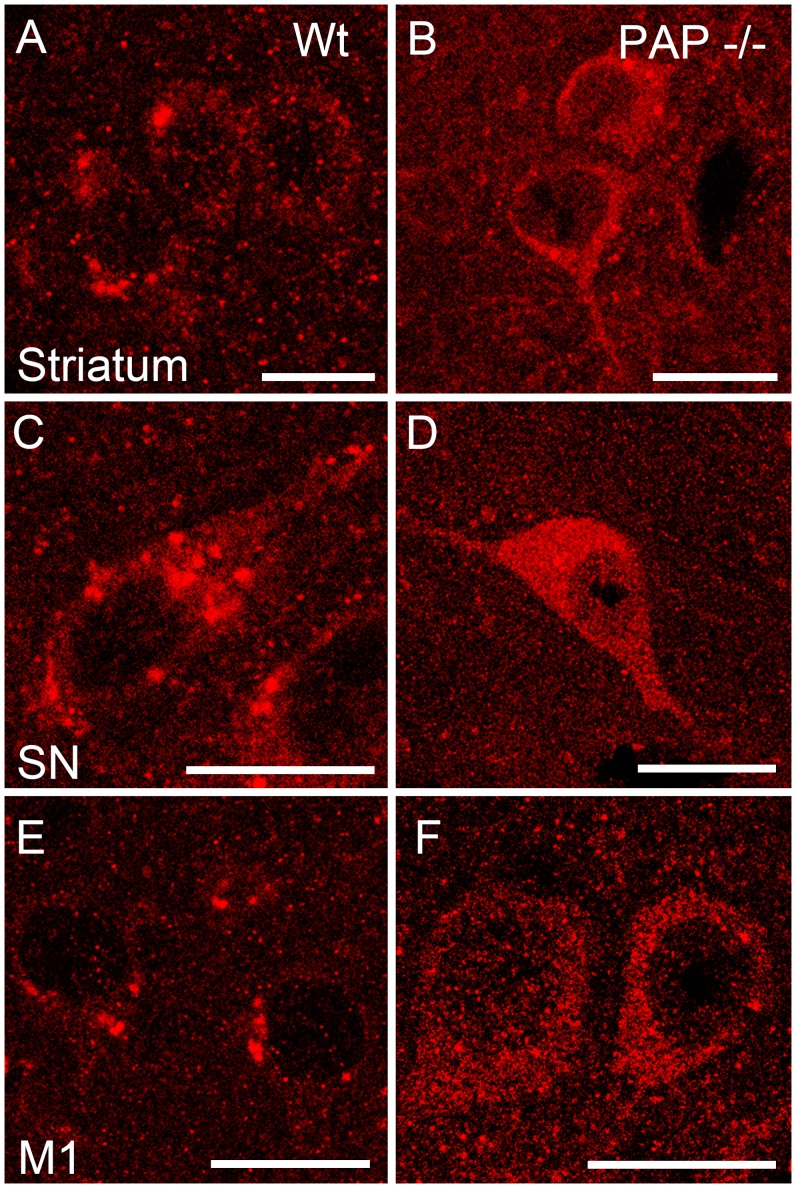
Prostatic acid phosphatase (PAP), the first diagnostic marker and present therapeutic target for prostate cancer, modulates nociception at the dorsal root ganglia (DRG), but its function in the central nervous system has remained unknown. We studied expression and function of TMPAP (the transmembrane isoform of PAP) in the brain by utilizing mice deficient in TMPAP (PAP-/- mice). Here we report that TMPAP is expressed in a subpopulation of cerebral GABAergic neurons, and mice deficient in TMPAP show multiple behavioral and neurochemical features linked to hyperdopaminergic dysregulation and altered GABAergic transmission. In addition to increased anxiety, disturbed prepulse inhibition, increased synthesis of striatal dopamine, and augmented response to amphetamine, PAP-deficient mice have enlarged lateral ventricles, reduced diazepam-induced loss of righting reflex, and increased GABAergic tone in the hippocampus. TMPAP in the mouse brain is localized presynaptically, and colocalized with SNARE-associated protein snapin, a protein involved in synaptic vesicle docking and fusion, and PAP-deficient mice display altered subcellular distribution of snapin. We have previously shown TMPAP to reside in prostatic exosomes and we propose that TMPAP is involved in the control of GABAergic tone in the brain also through exocytosis, and that PAP deficiency produces a distinct neurological phenotype.
Conflict of interest statement
Figures
References
-
- Quintero IB, Araujo CL, Pulkka AE, Wirkkala RS, Herrala AM, et al. (2007) Prostatic acid phosphatase is not a prostate specific target. Cancer Res 67: 6549–6554. - PubMed
-
- Vihko P (1979) Human prostatic acid phosphatases: Purification of a minor enzyme and comparisons of the enzymes. Invest Urol 16: 349–352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
