

The bicomponent pore-forming leucocidins of Staphylococcus aureus
- PMID: 24847020
- PMCID: PMC4054254
- DOI: 10.1128/MMBR.00055-13
The bicomponent pore-forming leucocidins of Staphylococcus aureus
Abstract
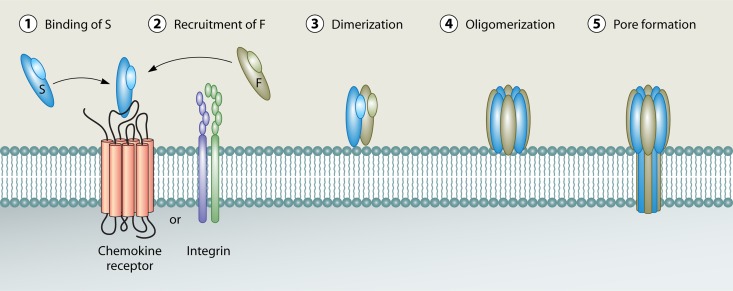
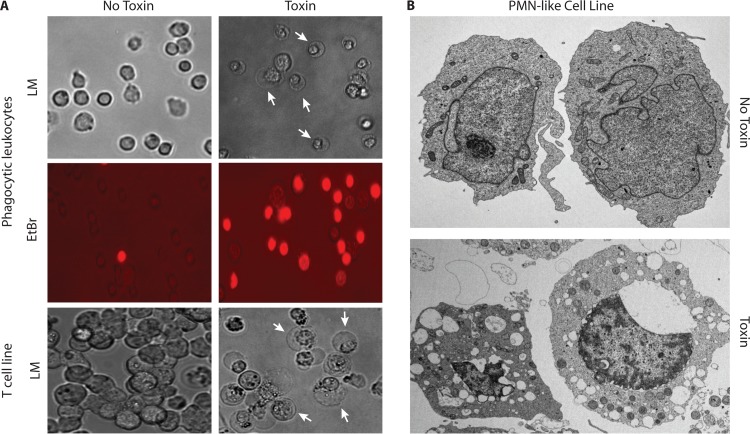
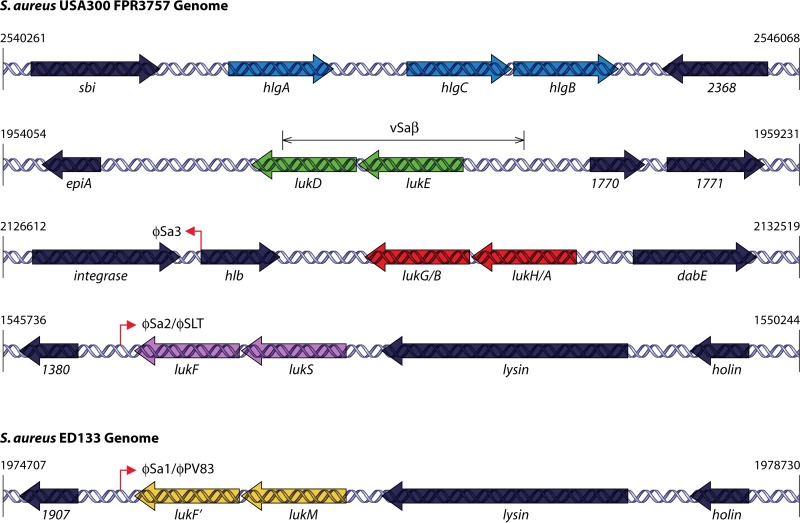
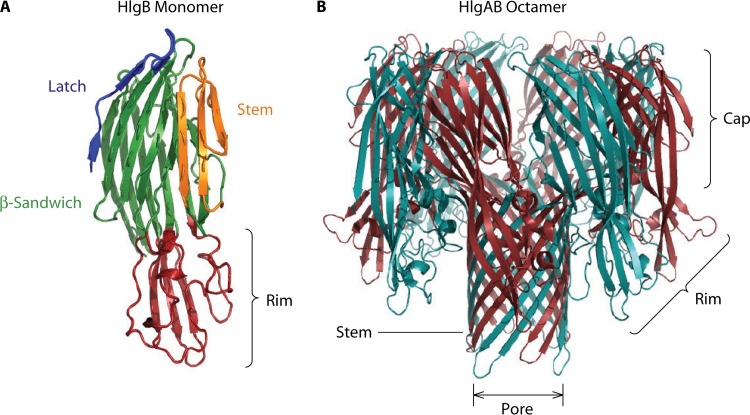
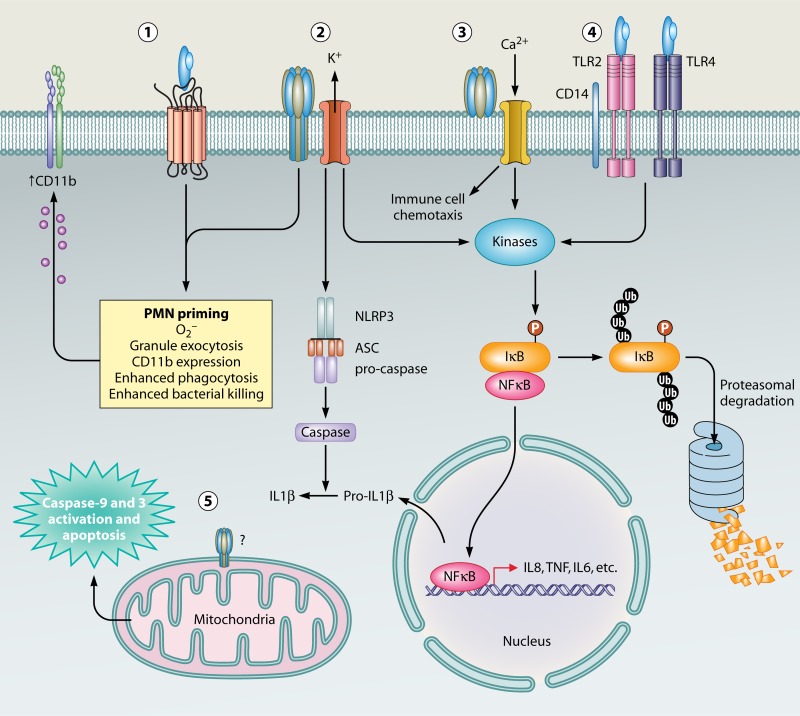
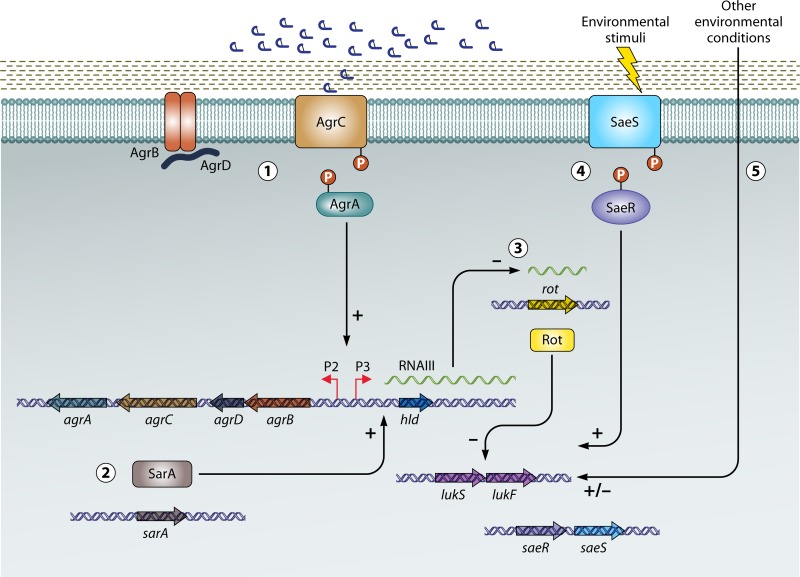
The ability to produce water-soluble proteins with the capacity to oligomerize and form pores within cellular lipid bilayers is a trait conserved among nearly all forms of life, including humans, single-celled eukaryotes, and numerous bacterial species. In bacteria, some of the most notable pore-forming molecules are protein toxins that interact with mammalian cell membranes to promote lysis, deliver effectors, and modulate cellular homeostasis. Of the bacterial species capable of producing pore-forming toxic molecules, the Gram-positive pathogen Staphylococcus aureus is one of the most notorious. S. aureus can produce seven different pore-forming protein toxins, all of which are believed to play a unique role in promoting the ability of the organism to cause disease in humans and other mammals. The most diverse of these pore-forming toxins, in terms of both functional activity and global representation within S. aureus clinical isolates, are the bicomponent leucocidins. From the first description of their activity on host immune cells over 100 years ago to the detailed investigations of their biochemical function today, the leucocidins remain at the forefront of S. aureus pathogenesis research initiatives. Study of their mode of action is of immediate interest in the realm of therapeutic agent design as well as for studies of bacterial pathogenesis. This review provides an updated perspective on our understanding of the S. aureus leucocidins and their function, specificity, and potential as therapeutic targets.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures



References
-
- Klevens RM, Morrison MA, Nadle J, Petit S, Gershman K, Ray S, Harrison LH, Lynfield R, Dumyati G, Townes JM, Craig AS, Zell ER, Fosheim GE, McDougal LK, Carey RB, Fridkin SK, Active Bacterial Core Surveillance MRSA Investigators 2007. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 298:1763–1771. 10.1001/jama.298.15.1763 - DOI - PubMed
-
- Klevens RM, Edwards JR, Tenover FC, McDonald LC, Horan T, Gaynes R, National Nosocomial Infections Surveillance System 2006. Changes in the epidemiology of methicillin-resistant Staphylococcus aureus in intensive care units in US hospitals, 1992-2003. Clin. Infect. Dis. 42:389–391. 10.1086/499367 - DOI - PubMed
-
- Liu C, Bayer A, Cosgrove SE, Daum RS, Fridkin SK, Gorwitz RJ, Kaplan SL, Karchmer AW, Levine DP, Murray BE, Ryback MJ, Talan DA, Chambers HF. 2011. Clinical practice guidelines by the Infectious Diseases Society of America for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children: executive summary. Clin. Infect. Dis. 52:285–292. 10.1093/cid/cir034 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
