

Impact of scaffold rigidity on the design and evolution of an artificial Diels-Alderase
- PMID: 24847076
- PMCID: PMC4050586
- DOI: 10.1073/pnas.1401073111
Impact of scaffold rigidity on the design and evolution of an artificial Diels-Alderase
Abstract
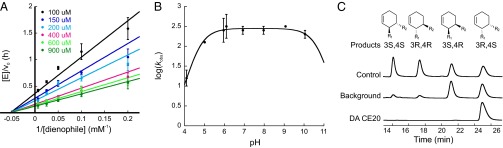
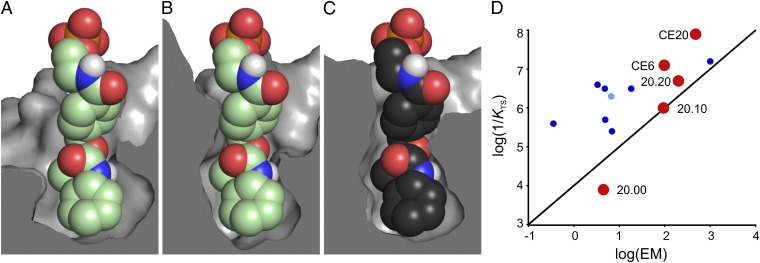
By combining targeted mutagenesis, computational refinement, and directed evolution, a modestly active, computationally designed Diels-Alderase was converted into the most proficient biocatalyst for [4+2] cycloadditions known. The high stereoselectivity and minimal product inhibition of the evolved enzyme enabled preparative scale synthesis of a single product diastereomer. X-ray crystallography of the enzyme-product complex shows that the molecular changes introduced over the course of optimization, including addition of a lid structure, gradually reshaped the pocket for more effective substrate preorganization and transition state stabilization. The good overall agreement between the experimental structure and the original design model with respect to the orientations of both the bound product and the catalytic side chains contrasts with other computationally designed enzymes. Because design accuracy appears to correlate with scaffold rigidity, improved control over backbone conformation will likely be the key to future efforts to design more efficient enzymes for diverse chemical reactions.
Keywords: Diels–Alder reaction; biocatalysis; computational enzyme design; enzyme mechanism; laboratory evolution.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Katayama K, Kobayashi T, Oikawa H, Honma M, Ichihara A. Enzymatic activity and partial purification of solanapyrone synthase: First enzyme catalyzing Diels-Alder reaction. Biochim Biophys Acta. 1998;1384(2):387–395. - PubMed
-
- Auclair K, et al. Lovastatin nonaketide synthase catalyzes an intramolecular Diels-Alder reaction of a substrate analogue. J Am Chem Soc. 2000;122(46):11519–11520.
-
- Stocking EM, Williams RM. Chemistry and biology of biosynthetic Diels-Alder reactions. Angew Chem Int Ed Engl. 2003;42(27):3078–3115. - PubMed
-
- Ose T, et al. Insight into a natural Diels-Alder reaction from the structure of macrophomate synthase. Nature. 2003;422(6928):185–189. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
