

Metformin suppresses gluconeogenesis by inhibiting mitochondrial glycerophosphate dehydrogenase
- PMID: 24847880
- PMCID: PMC4074244
- DOI: 10.1038/nature13270
Metformin suppresses gluconeogenesis by inhibiting mitochondrial glycerophosphate dehydrogenase
Abstract
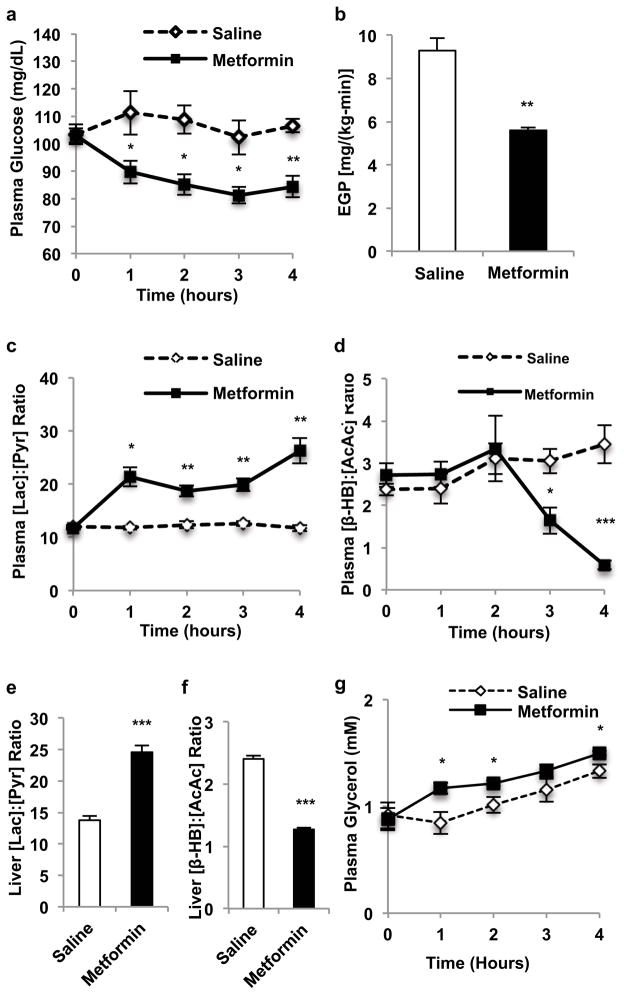
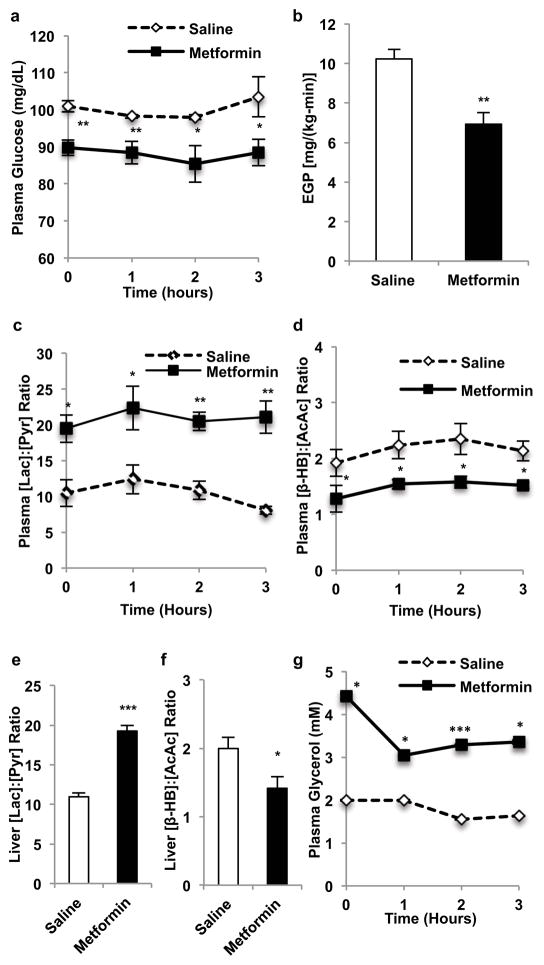
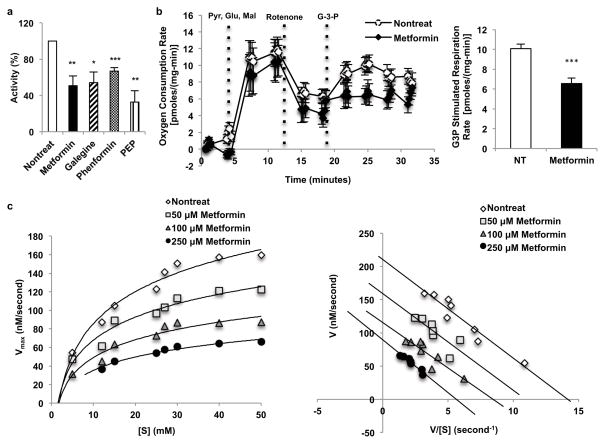
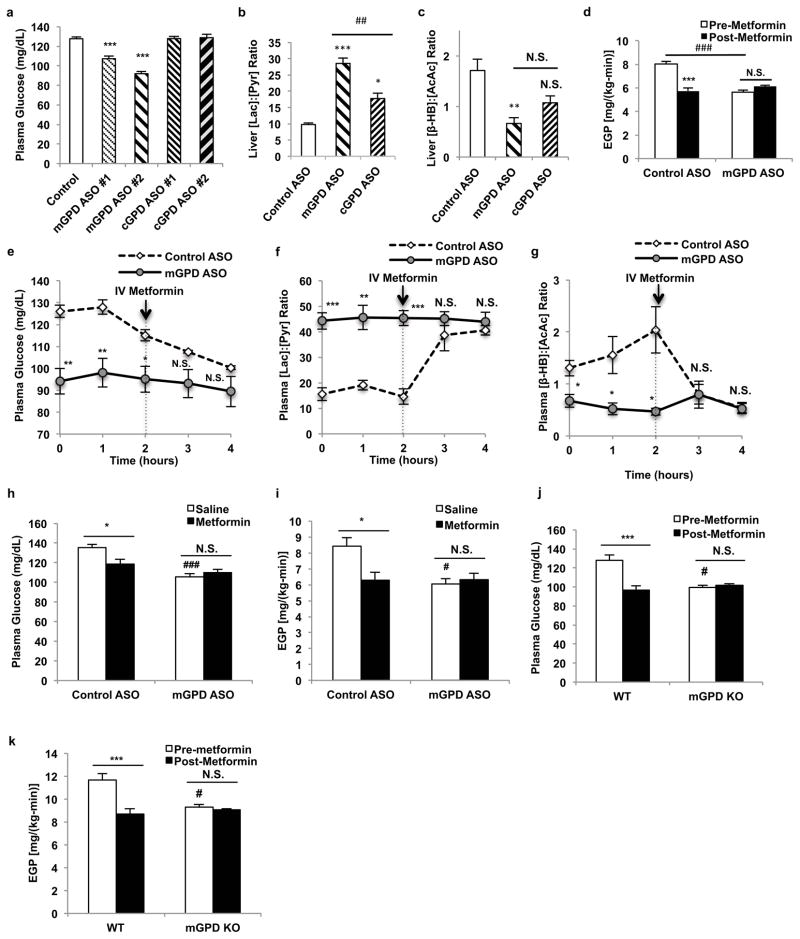
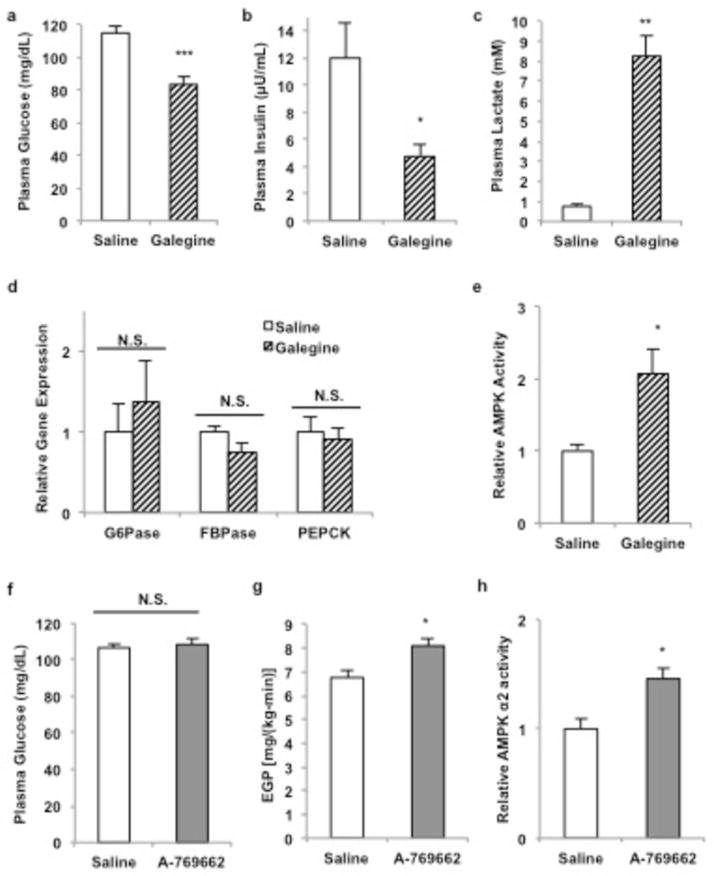
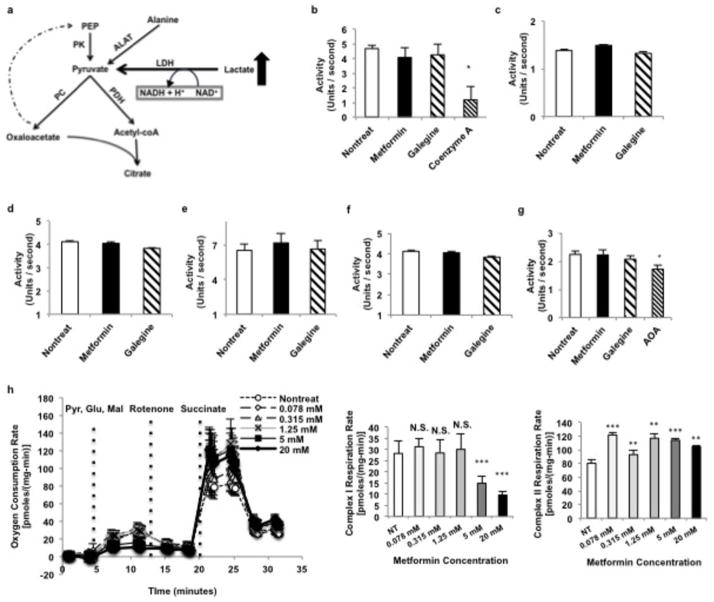
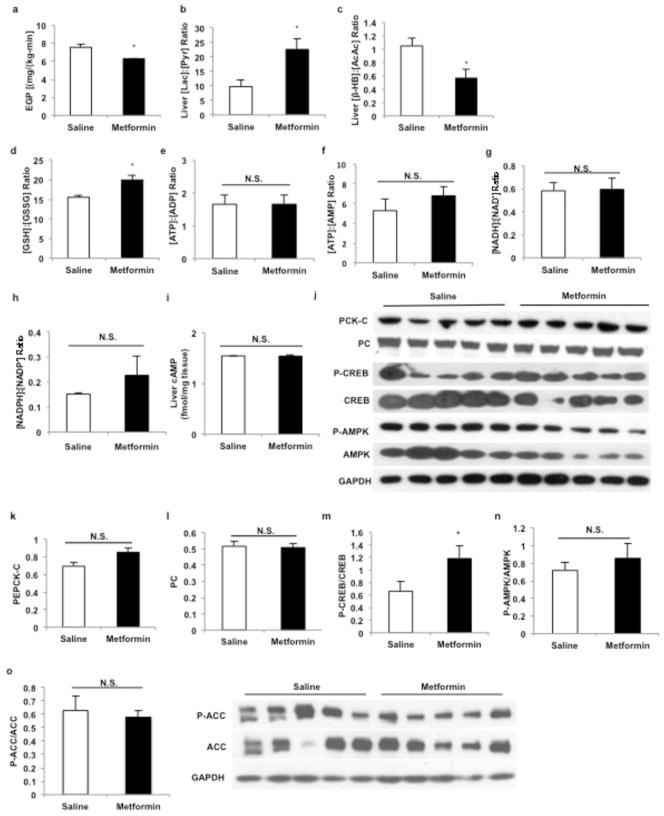
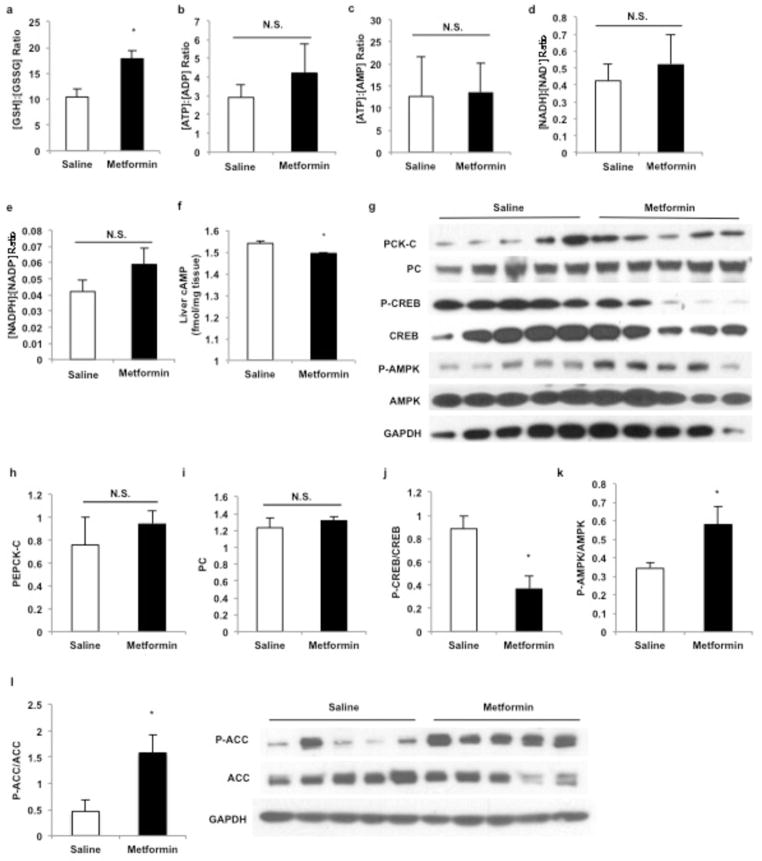
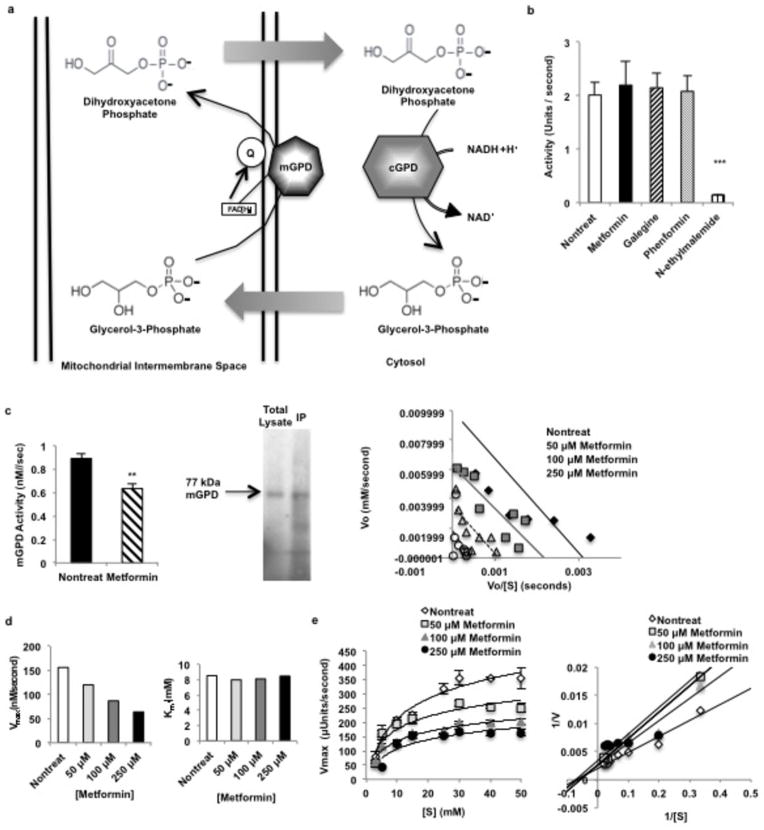
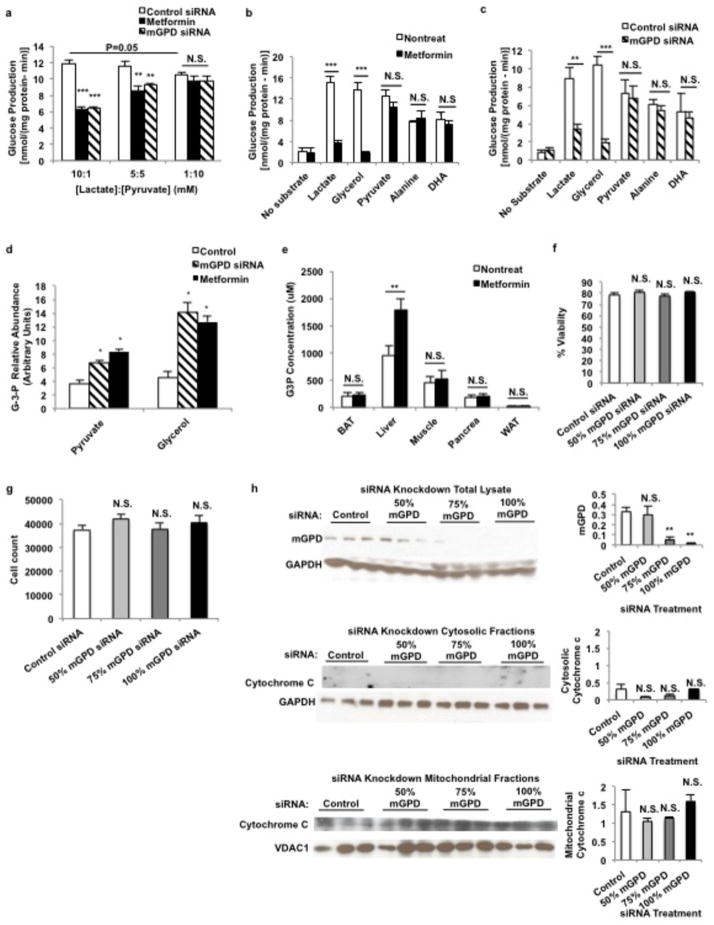
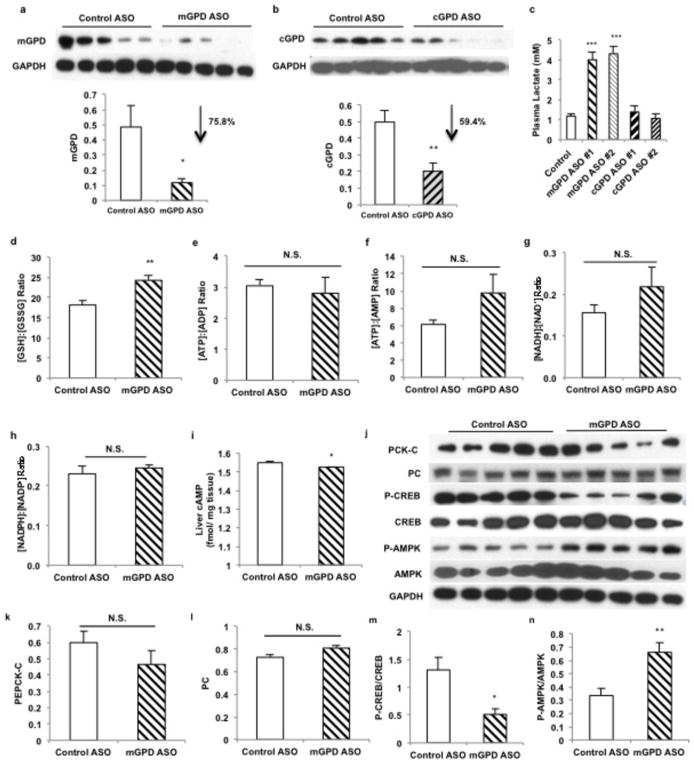
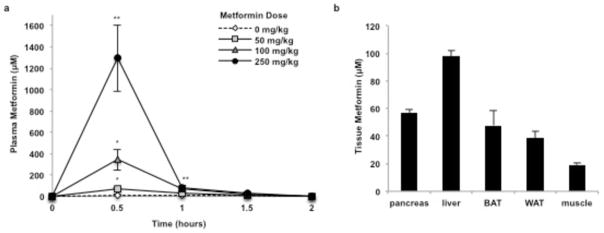
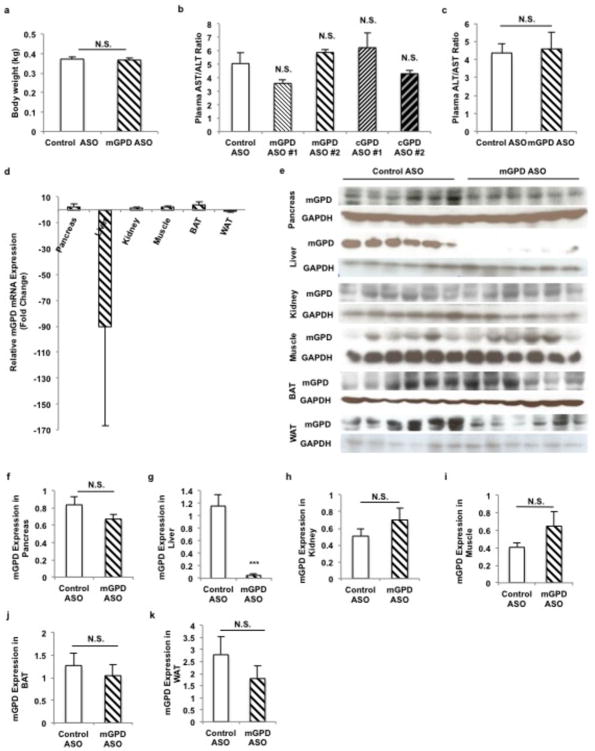
Metformin is considered to be one of the most effective therapeutics for treating type 2 diabetes because it specifically reduces hepatic gluconeogenesis without increasing insulin secretion, inducing weight gain or posing a risk of hypoglycaemia. For over half a century, this agent has been prescribed to patients with type 2 diabetes worldwide, yet the underlying mechanism by which metformin inhibits hepatic gluconeogenesis remains unknown. Here we show that metformin non-competitively inhibits the redox shuttle enzyme mitochondrial glycerophosphate dehydrogenase, resulting in an altered hepatocellular redox state, reduced conversion of lactate and glycerol to glucose, and decreased hepatic gluconeogenesis. Acute and chronic low-dose metformin treatment effectively reduced endogenous glucose production, while increasing cytosolic redox and decreasing mitochondrial redox states. Antisense oligonucleotide knockdown of hepatic mitochondrial glycerophosphate dehydrogenase in rats resulted in a phenotype akin to chronic metformin treatment, and abrogated metformin-mediated increases in cytosolic redox state, decreases in plasma glucose concentrations, and inhibition of endogenous glucose production. These findings were replicated in whole-body mitochondrial glycerophosphate dehydrogenase knockout mice. These results have significant implications for understanding the mechanism of metformin's blood glucose lowering effects and provide a new therapeutic target for type 2 diabetes.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Comment in
-
Control of gluconeogenesis by metformin: does redox trump energy charge?Cell Metab. 2014 Aug 5;20(2):197-9. doi: 10.1016/j.cmet.2014.07.013. Cell Metab. 2014. PMID: 25100057 Free PMC article.
References
-
- Inzucchi S, MD, Spollett GR, Page SL, Rife FS, Walton V, Shulman GI. Efficacy and metabolic effects of metformin and troglitazone in type II diabetes mellitus. N Eng J Med. 1998;338:867–872. - PubMed
-
- El-Mir M-Y, NV, Fontaine E, Averet N, Rigoulet M, Leverve X. Dimethylbiguanide Inhibits Cell Respiration via an Indirect Effect Targeted on the Respiratory Chain Complex I. J Biol Chem. 2000;275:223–228. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K01 DK099402/DK/NIDDK NIH HHS/United States
- R24 DK-085638/DK/NIDDK NIH HHS/United States
- R01 DK-40936/DK/NIDDK NIH HHS/United States
- R01 DK-092606/DK/NIDDK NIH HHS/United States
- P30 DK-45735/DK/NIDDK NIH HHS/United States
- R01 DK040936/DK/NIDDK NIH HHS/United States
- R24 DK085638/DK/NIDDK NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- UL1 TR000142/TR/NCATS NIH HHS/United States
- U24 DK-059635/DK/NIDDK NIH HHS/United States
- P30 DK-034989/DK/NIDDK NIH HHS/United States
- P30 DK034989/DK/NIDDK NIH HHS/United States
- R01 DK028348/DK/NIDDK NIH HHS/United States
- U24 DK059635/DK/NIDDK NIH HHS/United States
- K01 DK-099402/DK/NIDDK NIH HHS/United States
- P30 DK045735/DK/NIDDK NIH HHS/United States
- R01 DK-28348/DK/NIDDK NIH HHS/United States
- R01 DK092606/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
