

A cerebellar learning model of vestibulo-ocular reflex adaptation in wild-type and mutant mice
- PMID: 24849355
- PMCID: PMC6608186
- DOI: 10.1523/JNEUROSCI.2791-13.2014
A cerebellar learning model of vestibulo-ocular reflex adaptation in wild-type and mutant mice
Abstract
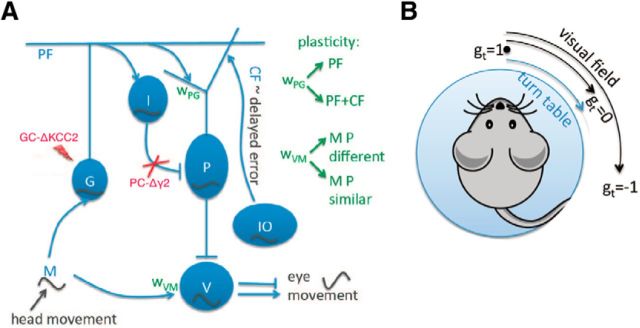
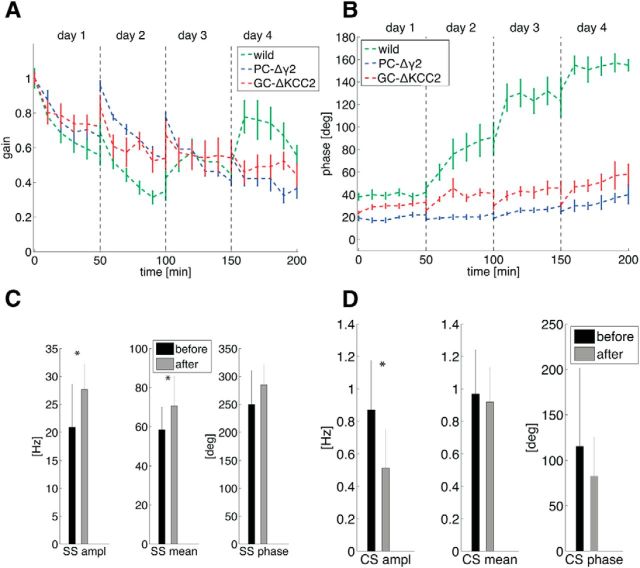
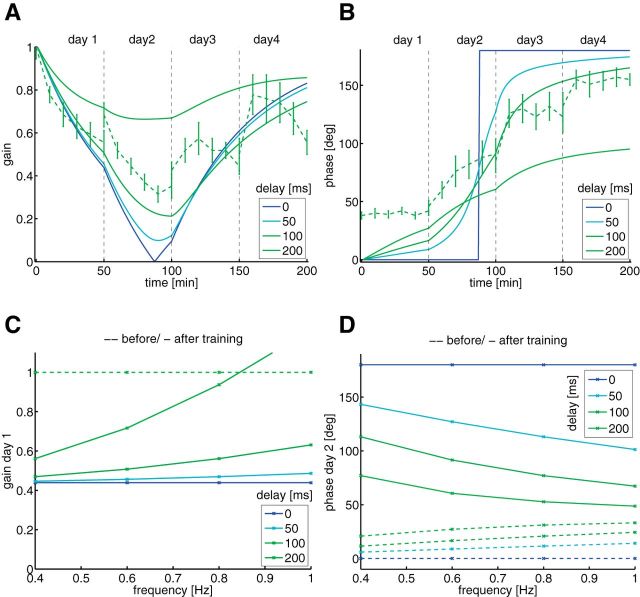
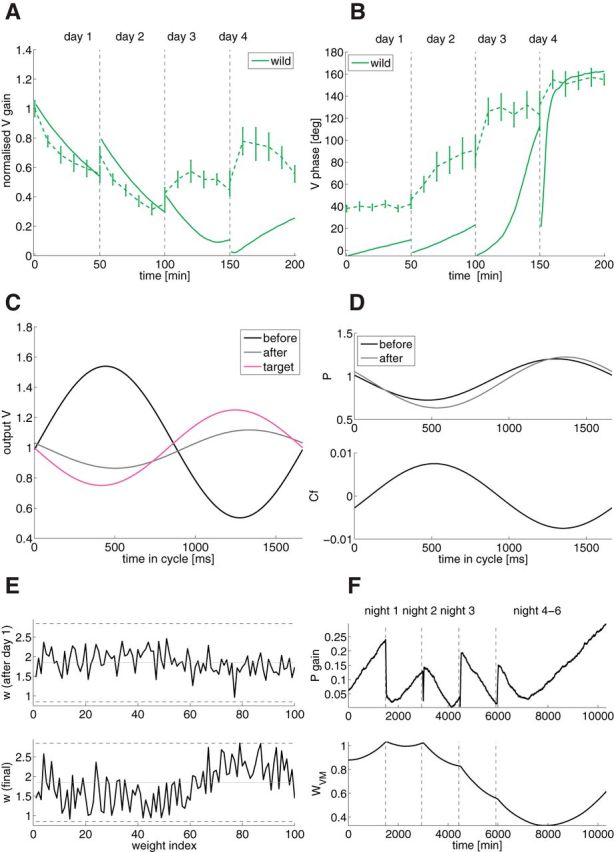
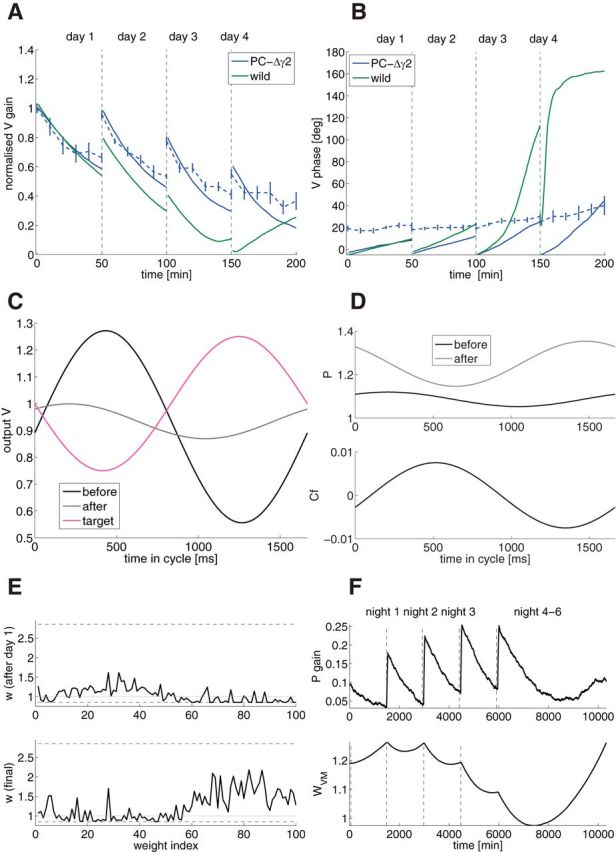
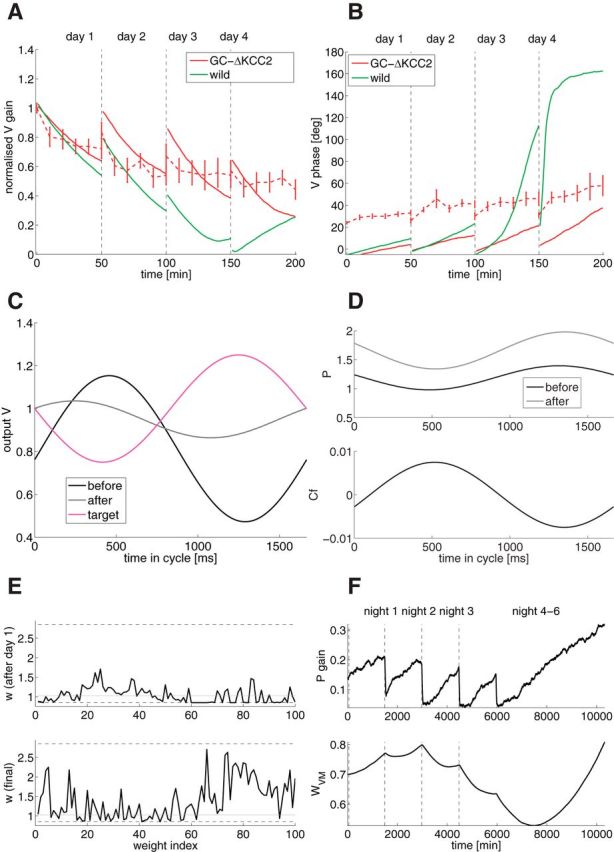
Mechanisms of cerebellar motor learning are still poorly understood. The standard Marr-Albus-Ito theory posits that learning involves plasticity at the parallel fiber to Purkinje cell synapses under control of the climbing fiber input, which provides an error signal as in classical supervised learning paradigms. However, a growing body of evidence challenges this theory, in that additional sites of plasticity appear to contribute to motor adaptation. Here, we consider phase-reversal training of the vestibulo-ocular reflex (VOR), a simple form of motor learning for which a large body of experimental data is available in wild-type and mutant mice, in which the excitability of granule cells or inhibition of Purkinje cells was affected in a cell-specific fashion. We present novel electrophysiological recordings of Purkinje cell activity measured in naive wild-type mice subjected to this VOR adaptation task. We then introduce a minimal model that consists of learning at the parallel fibers to Purkinje cells with the help of the climbing fibers. Although the minimal model reproduces the behavior of the wild-type animals and is analytically tractable, it fails at reproducing the behavior of mutant mice and the electrophysiology data. Therefore, we build a detailed model involving plasticity at the parallel fibers to Purkinje cells' synapse guided by climbing fibers, feedforward inhibition of Purkinje cells, and plasticity at the mossy fiber to vestibular nuclei neuron synapse. The detailed model reproduces both the behavioral and electrophysiological data of both the wild-type and mutant mice and allows for experimentally testable predictions.
Copyright © 2014 the authors 0270-6474/14/347203-13$15.00/0.
Figures
References
-
- Albus JS. A theory of cerebellar function. Math Biosci. 1971;10:25–61. doi: 10.1016/0025-5564(71)90051-4. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases