

Feeling the hidden mechanical forces in lipid bilayer is an original sense
- PMID: 24850861
- PMCID: PMC4050596
- DOI: 10.1073/pnas.1313364111
Feeling the hidden mechanical forces in lipid bilayer is an original sense
Abstract
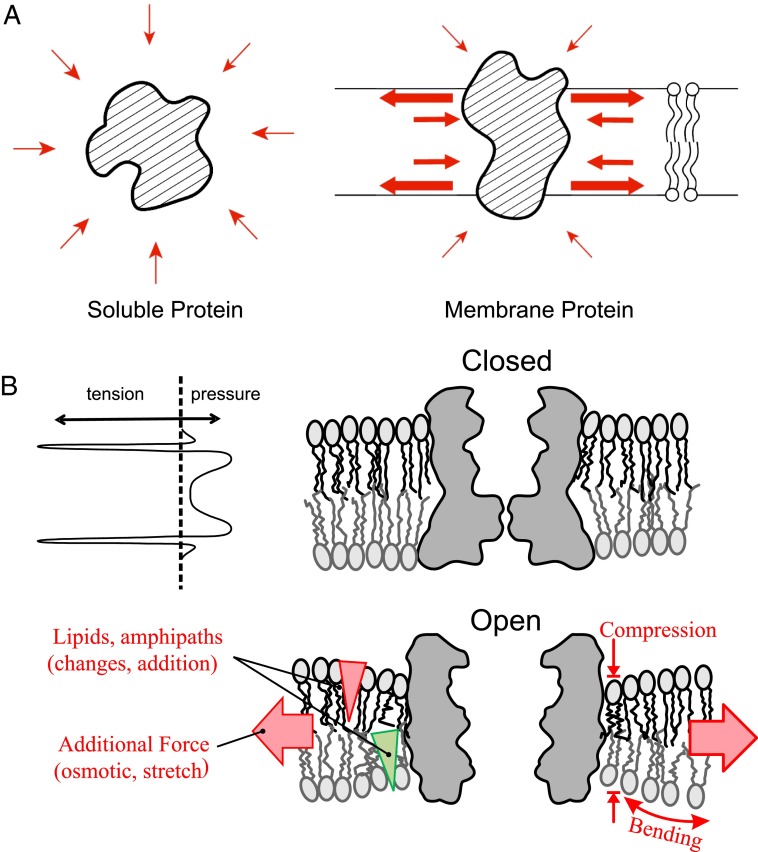
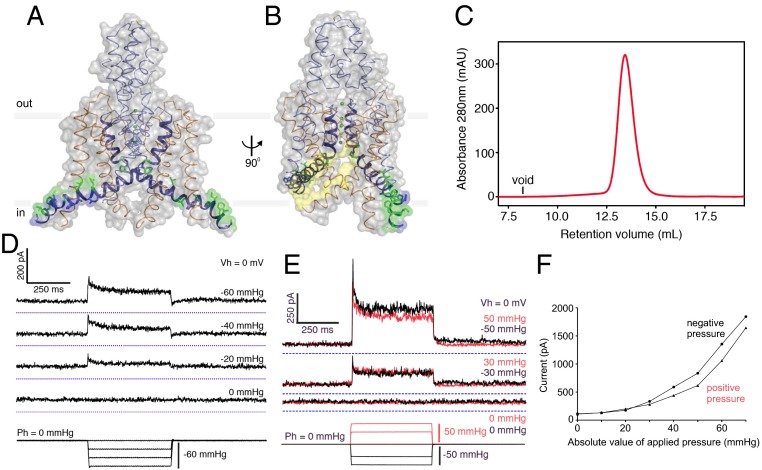
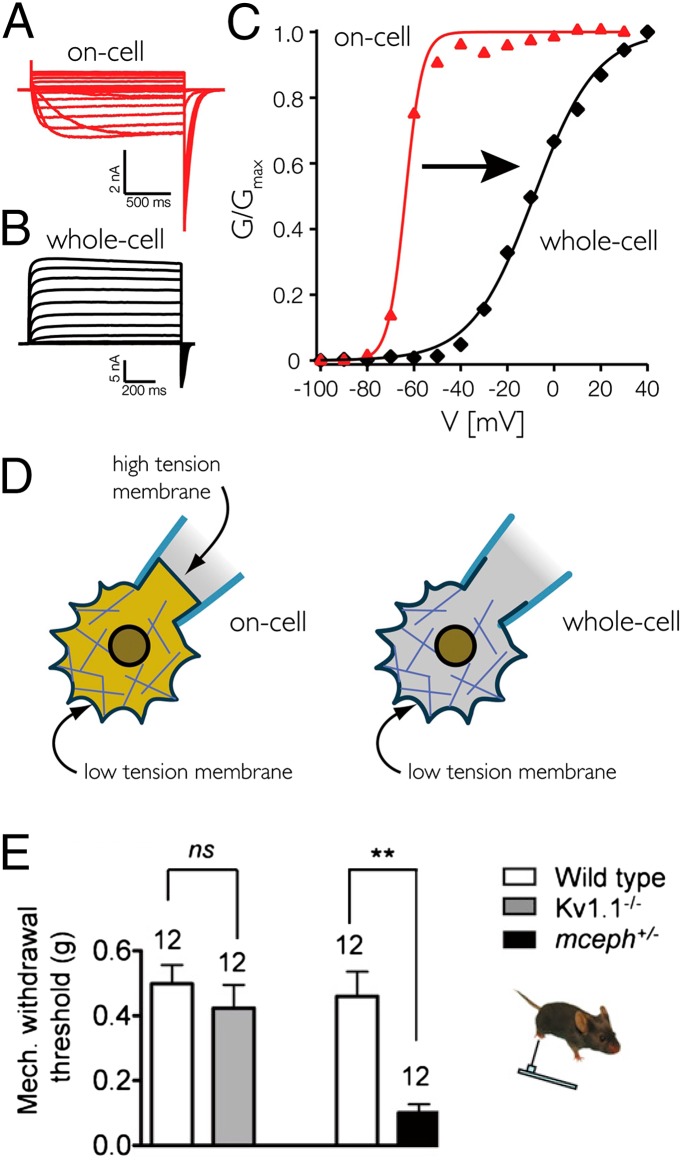
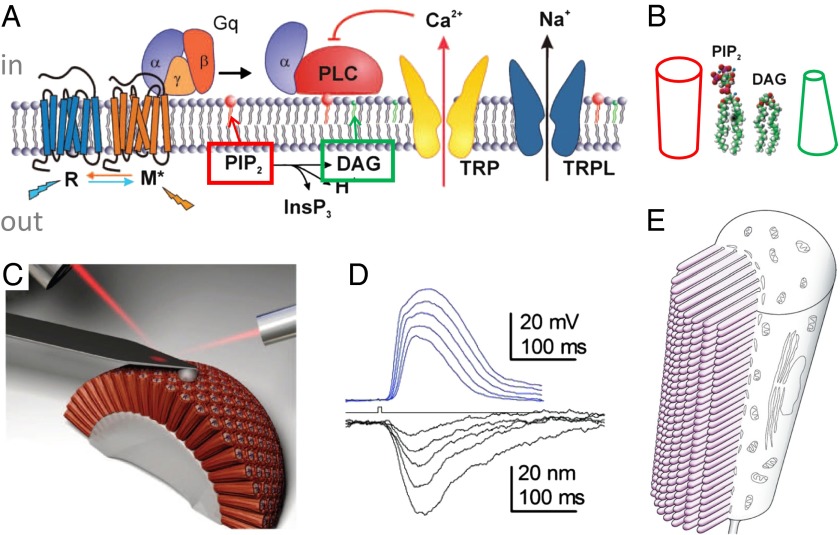
Life's origin entails enclosing a compartment to hoard material, energy, and information. The envelope necessarily comprises amphipaths, such as prebiotic fatty acids, to partition the two aqueous domains. The self-assembled lipid bilayer comes with a set of properties including its strong anisotropic internal forces that are chemically or physically malleable. Added bilayer stretch can alter force vectors on embedded proteins to effect conformational change. The force-from-lipid principle was demonstrated 25 y ago when stretches opened purified Escherichia coli MscL channels reconstituted into artificial bilayers. This reductionistic exercise has rigorously been recapitulated recently with two vertebrate mechanosensitive K(+) channels (TREK1 and TRAAK). Membrane stretches have also been known to activate various voltage-, ligand-, or Ca(2+)-gated channels. Careful analyses showed that Kv, the canonical voltage-gated channel, is in fact exquisitely sensitive even to very small tension. In an unexpected context, the canonical transient-receptor-potential channels in the Drosophila eye, long presumed to open by ligand binding, is apparently opened by membrane force due to PIP2 hydrolysis-induced changes in bilayer strain. Being the intimate medium, lipids govern membrane proteins by physics as well as chemistry. This principle should not be a surprise because it parallels water's paramount role in the structure and function of soluble proteins. Today, overt or covert mechanical forces govern cell biological processes and produce sensations. At the genesis, a bilayer's response to osmotic force is likely among the first senses to deal with the capricious primordial sea.
Keywords: bilayer mechanics; channel gating; force sensing; mechanosensitivity; touch.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Long SB, Campbell EB, Mackinnon R. Crystal structure of a mammalian voltage-dependent Shaker family K+ channel. Science. 2005;309(5736):897–903. - PubMed
-
- Beyer MK, Clausen-Schaumann H. Mechanochemistry: The mechanical activation of covalent bonds. Chem Rev. 2005;105(8):2921–2948. - PubMed
-
- Wang W, Donini O, Reyes CM, Kollman PA. Biomolecular simulations: Recent developments in force fields, simulations of enzyme catalysis, protein-ligand, protein-protein, and protein-nucleic acid noncovalent interactions. Annu Rev Biophys Biomol Struct. 2001;30:211–243. - PubMed
-
- Khalid S, Bond PJ. Multiscale molecular dynamics simulations of membrane proteins. Methods Mol Biol. 2013;924:635–657. - PubMed
-
- Stansfeld PJ, Sansom MS. Molecular simulation approaches to membrane proteins. Structure. 2011;19(11):1562–1572. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
