

Molecular-crowding effects on single-molecule RNA folding/unfolding thermodynamics and kinetics
- PMID: 24850865
- PMCID: PMC4060727
- DOI: 10.1073/pnas.1316039111
Molecular-crowding effects on single-molecule RNA folding/unfolding thermodynamics and kinetics
Abstract
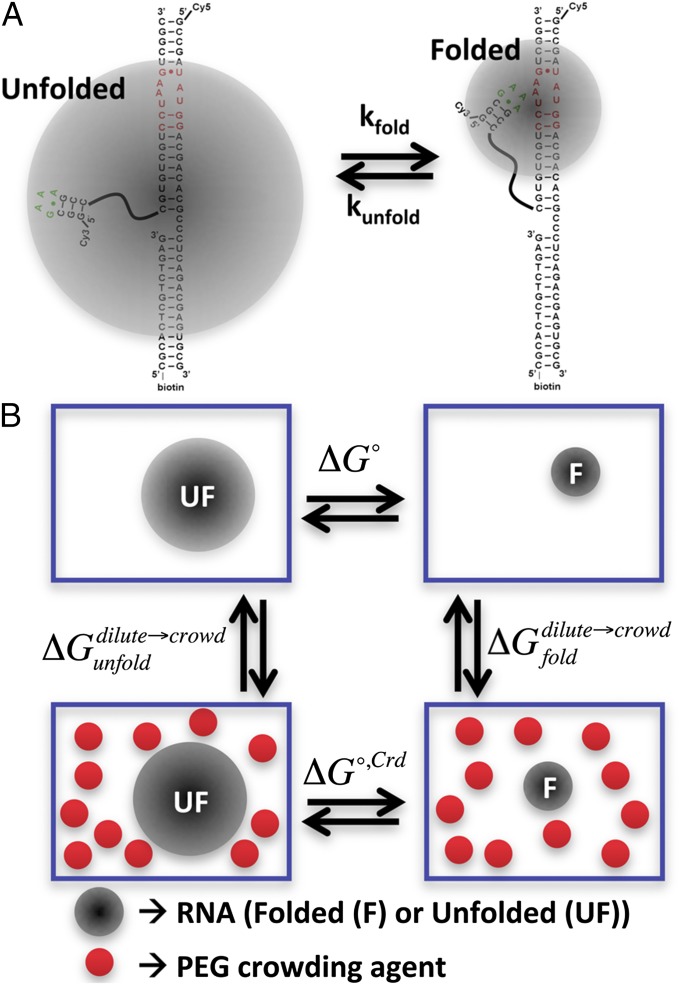
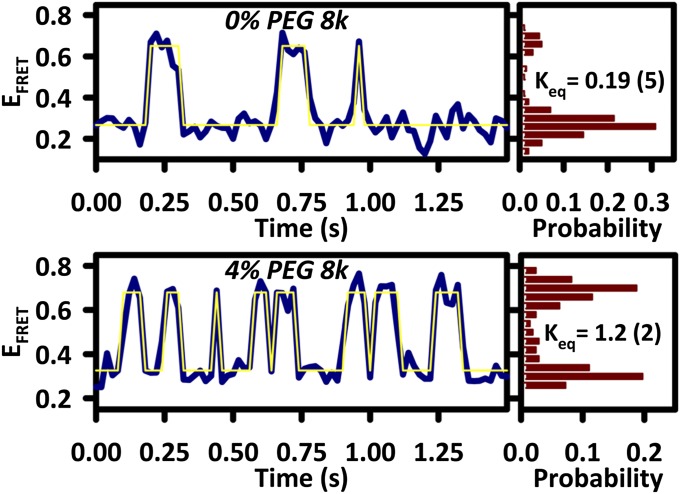
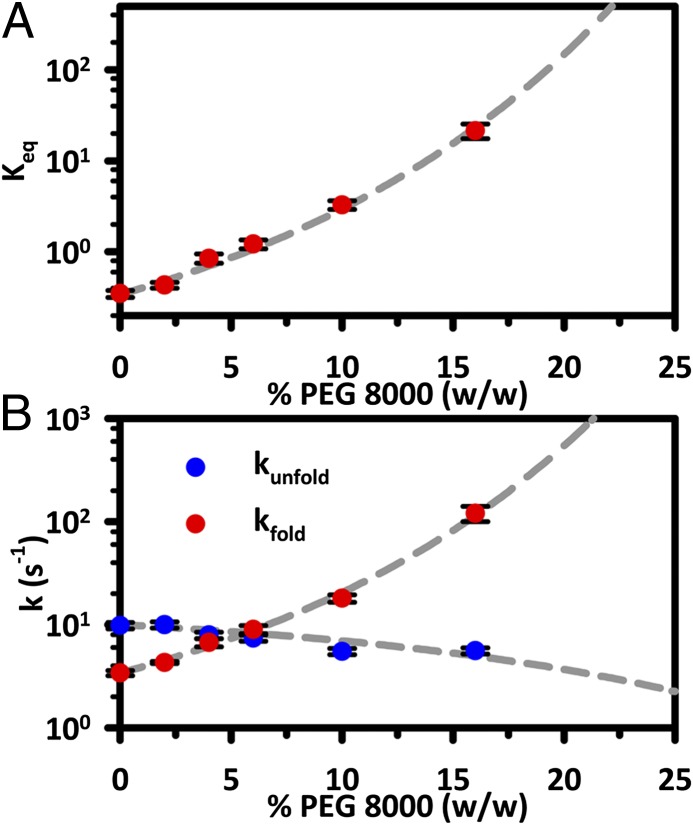
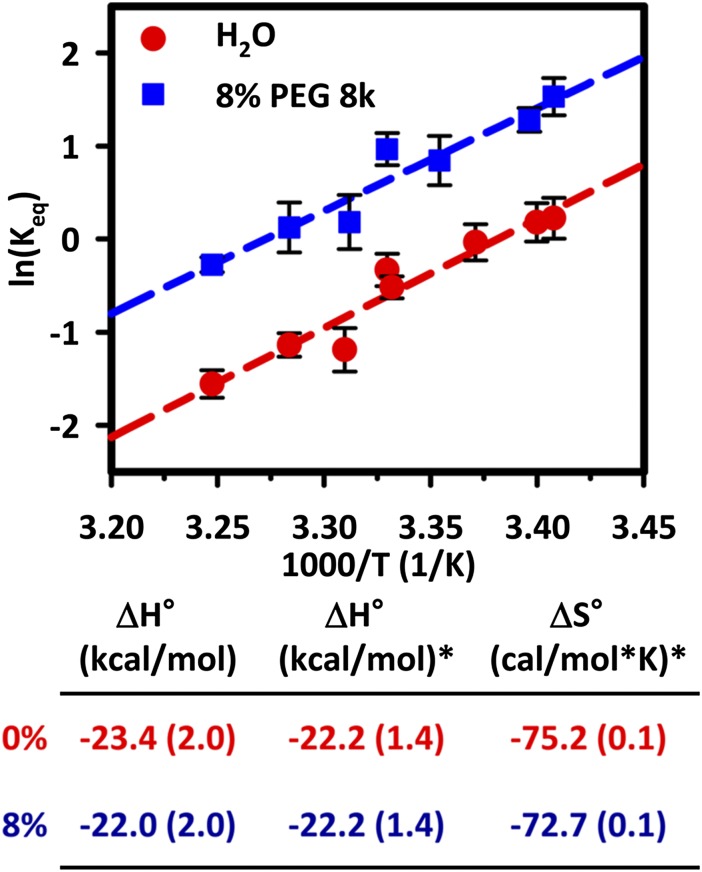
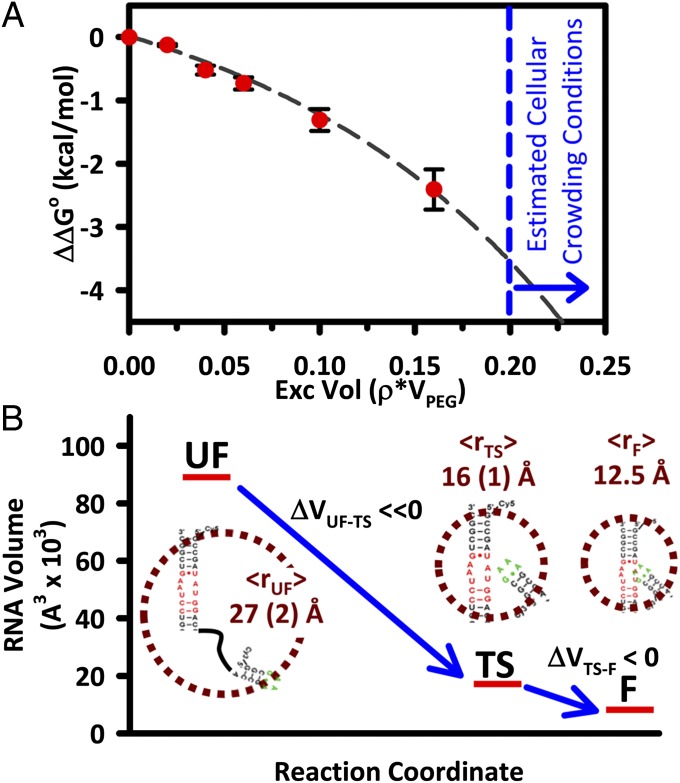
The effects of "molecular crowding" on elementary biochemical processes due to high solute concentrations are poorly understood and yet clearly essential to the folding of nucleic acids and proteins into correct, native structures. The present work presents, to our knowledge, first results on the single-molecule kinetics of solute molecular crowding, specifically focusing on GAAA tetraloop-receptor folding to isolate a single RNA tertiary interaction using time-correlated single-photon counting and confocal single-molecule FRET microscopy. The impact of crowding by high-molecular-weight polyethylene glycol on the RNA folding thermodynamics is dramatic, with up to ΔΔG° ∼ -2.5 kcal/mol changes in free energy and thus >60-fold increase in the folding equilibrium constant (Keq) for excluded volume fractions of 15%. Most importantly, time-correlated single-molecule methods permit crowding effects on the kinetics of RNA folding/unfolding to be explored for the first time (to our knowledge), which reveal that this large jump in Keq is dominated by a 35-fold increase in tetraloop-receptor folding rate, with only a modest decrease in the corresponding unfolding rate. This is further explored with temperature-dependent single-molecule RNA folding measurements, which identify that crowding effects are dominated by entropic rather than enthalpic contributions to the overall free energy change. Finally, a simple "hard-sphere" treatment of the solute excluded volume is invoked to model the observed kinetic trends, and which predict ΔΔG° ∼ -5 kcal/mol free-energy stabilization at excluded volume fractions of 30%.
Keywords: PEG; fluorescence; scaled particle theory.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Atkins JF, Gesteland RF, Cech T. RNA Worlds: From Life's Origins to Diversity in Gene Regulation. Cold Spring Harbor, NY: Cold Spring Harbor Lab Press; 2011.
-
- Butcher SE, Pyle AM. The molecular interactions that stabilize RNA tertiary structure: RNA motifs, patterns, and networks. Acc Chem Res. 2011;44(12):1302–1311. - PubMed
-
- Brion P, Westhof E. Hierarchy and dynamics of RNA folding. Annu Rev Biophys Biomol Struct. 1997;26:113–137. - PubMed
-
- Sear RP. The cytoplasm of living cells: A functional mixture of thousands of components. J Phys Condens Matter. 2005;17(45):S3587–S3595.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
