

None of the rotor residues of F1-ATPase are essential for torque generation
- PMID: 24853745
- PMCID: PMC4052266
- DOI: 10.1016/j.bpj.2014.04.013
None of the rotor residues of F1-ATPase are essential for torque generation
Abstract
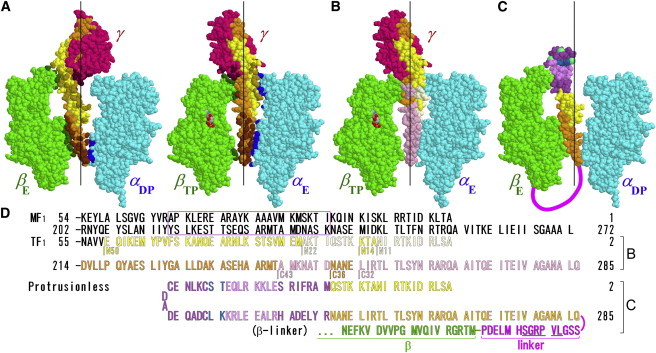
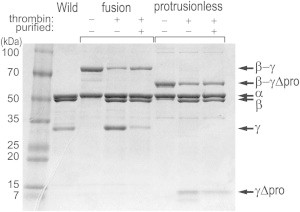
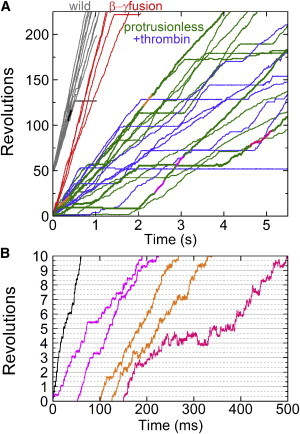
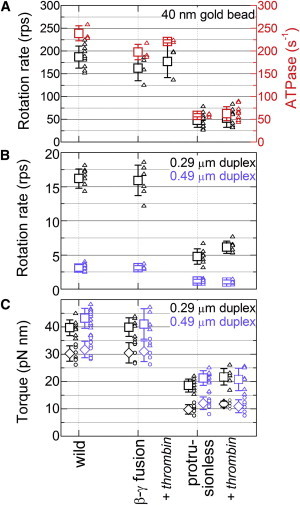
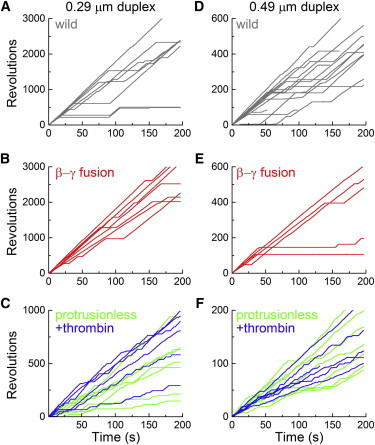
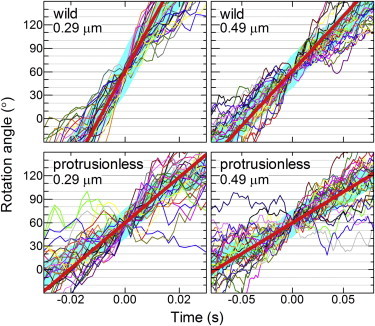
F1-ATPase is a powerful rotary molecular motor that can rotate an object several hundred times as large as the motor itself against the viscous friction of water. Forced reverse rotation has been shown to lead to ATP synthesis, implying that the mechanical work against the motor's high torque can be converted into the chemical energy of ATP. The minimal composition of the motor protein is α3β3γ subunits, where the central rotor subunit γ turns inside a stator cylinder made of alternately arranged α3β3 subunits using the energy derived from ATP hydrolysis. The rotor consists of an axle, a coiled coil of the amino- and carboxyl-terminal α-helices of γ, which deeply penetrates the stator cylinder, and a globular protrusion that juts out from the stator. Previous work has shown that, for a thermophilic F1, significant portions of the axle can be truncated and the motor still rotates a submicron sized bead duplex, indicating generation of up to half the wild-type (WT) torque. Here, we inquire if any specific interactions between the stator and the rest of the rotor are needed for the generation of a sizable torque. We truncated the protruding portion of the rotor and replaced part of the remaining axle residues such that every residue of the rotor has been deleted or replaced in this or previous truncation mutants. This protrusionless construct showed an unloaded rotary speed about a quarter of the WT, and generated one-third to one-half of the WT torque. No residue-specific interactions are needed for this much performance. F1 is so designed that the basic rotor-stator interactions for torque generation and control of catalysis rely solely upon the shape and size of the rotor at very low resolution. Additional tailored interactions augment the torque to allow ATP synthesis under physiological conditions.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Noji H., Yasuda R., Kinosita K., Jr. Direct observation of the rotation of F1-ATPase. Nature. 1997;386:299–302. - PubMed
-
- Yoshida M., Muneyuki E., Hisabori T. ATP synthase—a marvellous rotary engine of the cell. Nat. Rev. Mol. Cell Biol. 2001;2:669–677. - PubMed
-
- Boyer P.D. A research journey with ATP synthase. J. Biol. Chem. 2002;277:39045–39061. - PubMed
-
- Senior A.E., Nadanaciva S., Weber J. The molecular mechanism of ATP synthesis by F1F0-ATP synthase. Biochim. Biophys. Acta. 2002;1553:188–211. - PubMed
-
- Kinosita K., Jr., Adachi K., Itoh H. Rotation of F1-ATPase: how an ATP-driven molecular machine may work. Annu. Rev. Biophys. Biomol. Struct. 2004;33:245–268. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
