

Compensatory flux changes within an endocytic trafficking network maintain thermal robustness of Notch signaling
- PMID: 24855951
- PMCID: PMC4032575
- DOI: 10.1016/j.cell.2014.03.050
Compensatory flux changes within an endocytic trafficking network maintain thermal robustness of Notch signaling
Abstract
Developmental signaling is remarkably robust to environmental variation, including temperature. For example, in ectothermic animals such as Drosophila, Notch signaling is maintained within functional limits across a wide temperature range. We combine experimental and computational approaches to show that temperature compensation of Notch signaling is achieved by an unexpected variety of endocytic-dependent routes to Notch activation which, when superimposed on ligand-induced activation, act as a robustness module. Thermal compensation arises through an altered balance of fluxes within competing trafficking routes, coupled with temperature-dependent ubiquitination of Notch. This flexible ensemble of trafficking routes supports Notch signaling at low temperature but can be switched to restrain Notch signaling at high temperature and thus compensates for the inherent temperature sensitivity of ligand-induced activation. The outcome is to extend the physiological range over which normal development can occur. Similar mechanisms may provide thermal robustness for other developmental signals.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures

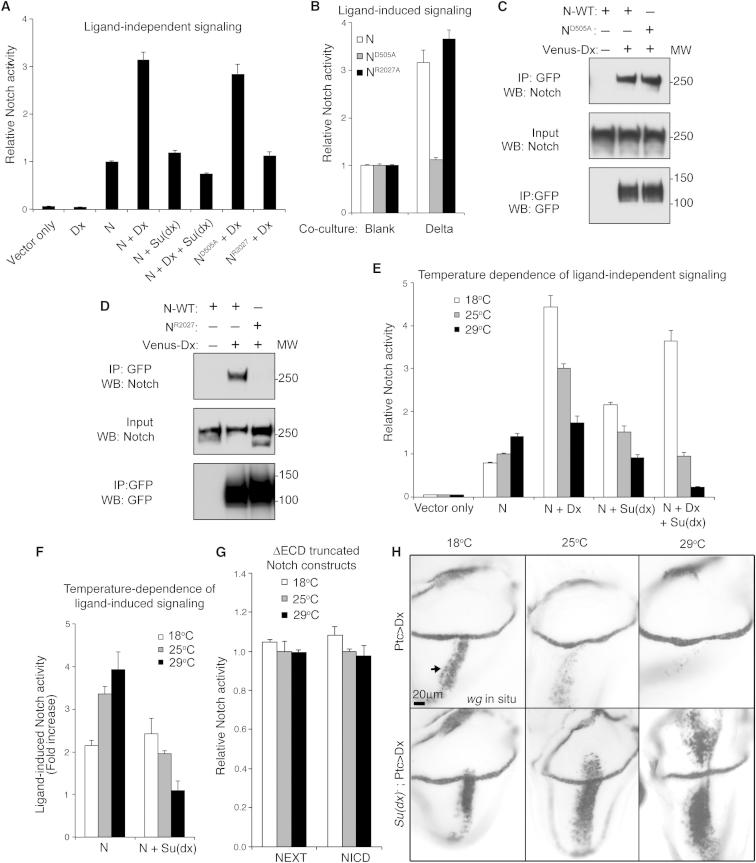
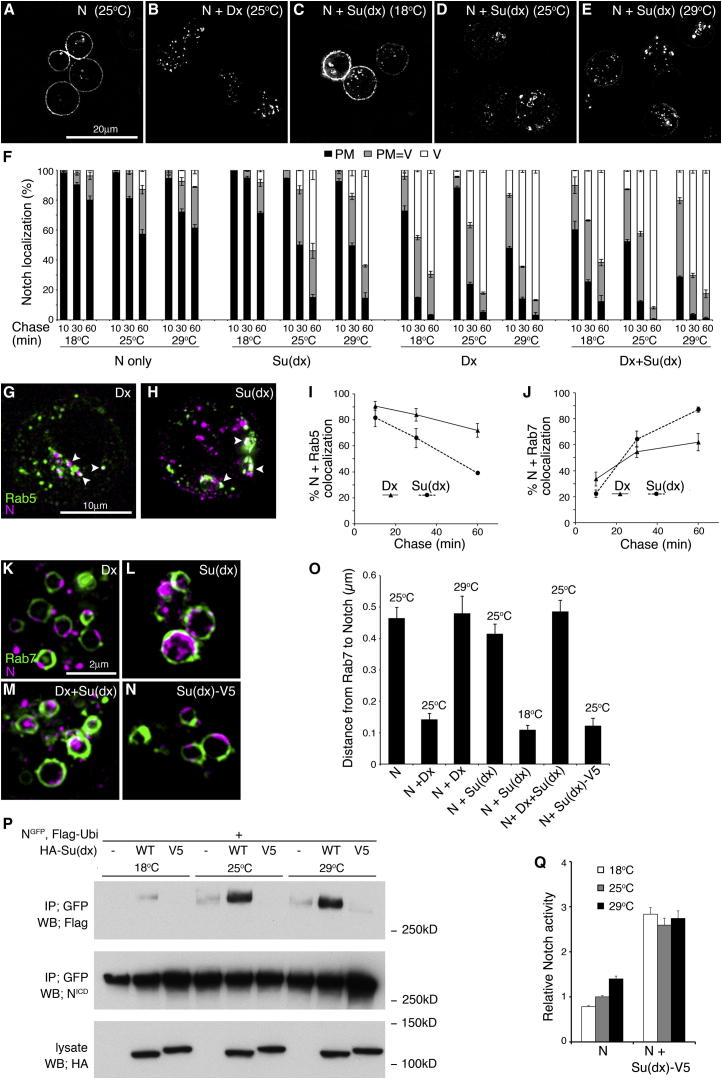
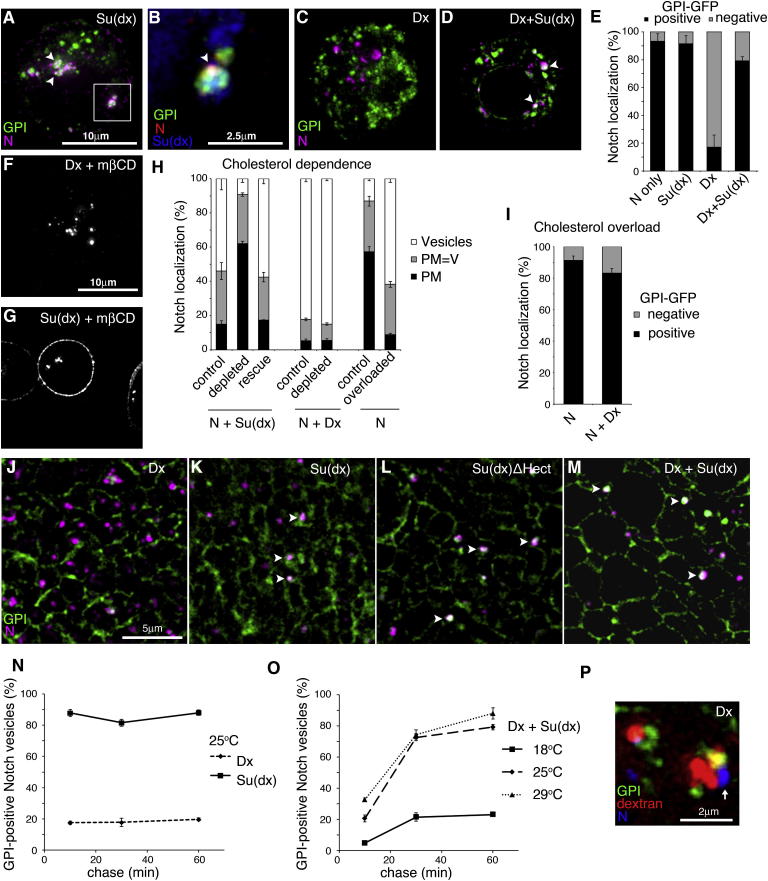


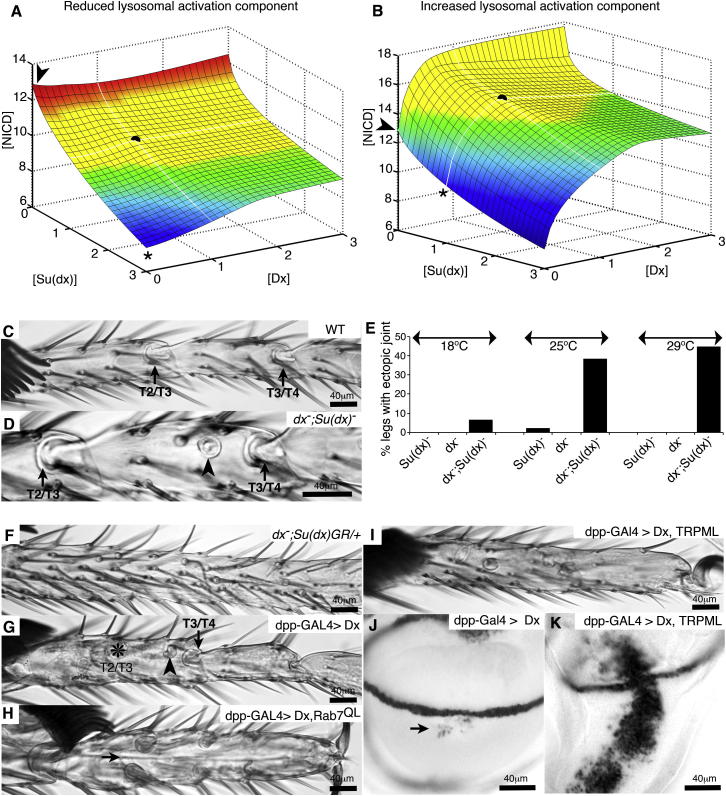
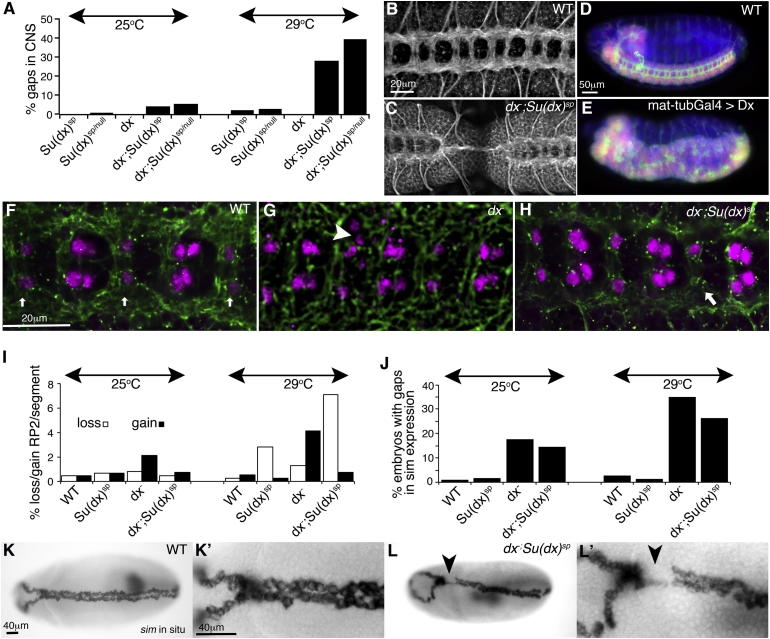

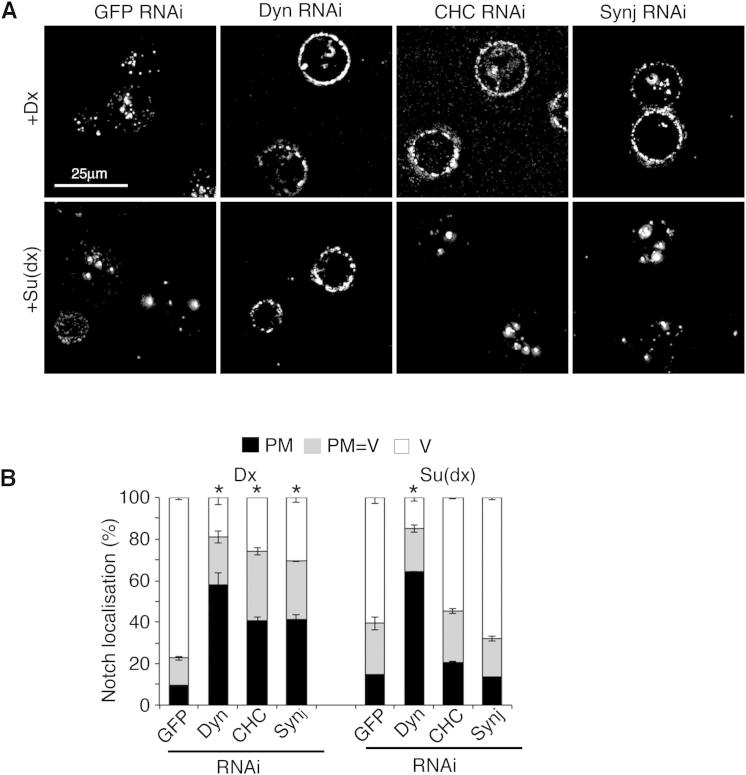

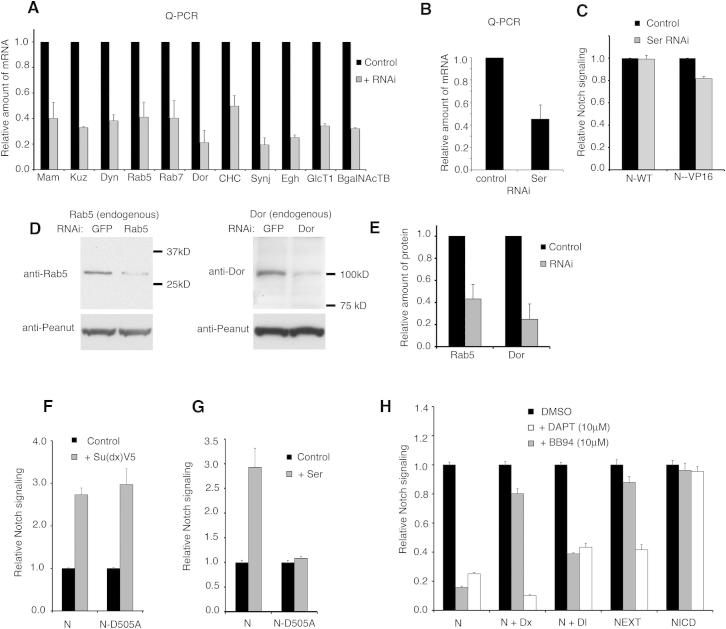
Comment in
-
Tempering temperature changes for robust development.Cell. 2014 May 22;157(5):1010-2. doi: 10.1016/j.cell.2014.05.007. Cell. 2014. PMID: 24855940
References
-
- Baron M. Endocytic routes to Notch activation. Semin. Cell Dev. Biol. 2012;23:437–442. - PubMed
-
- Bishop S.A., Klein T., Arias A.M., Couso J.P. Composite signalling from Serrate and Delta establishes leg segments in Drosophila through Notch. Development. 1999;126:2993–3003. - PubMed
-
- Christian A.E., Haynes M.P., Phillips M.C., Rothblat G.H. Use of cyclodextrins for manipulating cellular cholesterol content. J. Lipid Res. 1997;38:2264–2272. - PubMed
Supplemental References
-
- Diederich R.J., Matsuno K., Hing H., Artavanis-Tsakonas S. Cytosolic interaction between deltex and Notch ankyrin repeats implicates deltex in the Notch signaling pathway. Development. 1994;120:473–481. - PubMed
-
- Entchev E.V., Schwabedissen A., González-Gaitán M. Gradient formation of the TGF-beta homolog Dpp. Cell. 2000;103:981–991. - PubMed
-
- Flasza M., Nguyen Huu N.S., Mazaleyrat S., Clémence S., Villemant C., Clarke R., Baron M. Regulation of the nuclear localization of the human Nedd4-related WWP1 protein by Notch. Mol. Membr. Biol. 2006;23:269–276. - PubMed
-
- Fuwa T.J., Hori K., Sasamura T., Higgs J., Baron M., Matsuno K. The first deltex null mutant indicates tissue-specific deltex-dependent Notch signaling in Drosophila. Mol. Genet. Genomics. 2006;275:251–263. - PubMed
-
- Greco V., Hannus M., Eaton S. Argosomes: a potential vehicle for the spread of morphogens through epithelia. Cell. 2001;106:633–645. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/E002285/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/C503162/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- H000976/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 087928/WT_/Wellcome Trust/United Kingdom
- WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
