

MicroRNA-directed cleavage of targets: mechanism and experimental approaches
- PMID: 24856832
- PMCID: PMC4206712
- DOI: 10.5483/bmbrep.2014.47.8.109
MicroRNA-directed cleavage of targets: mechanism and experimental approaches
Abstract
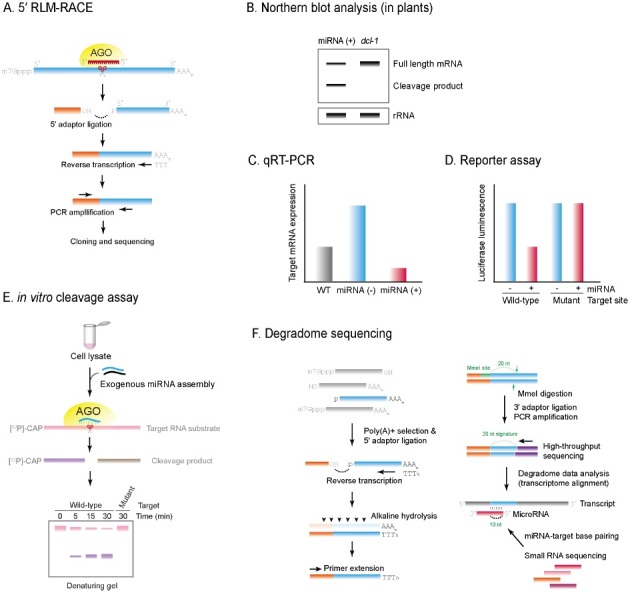
MicroRNAs (miRNAs) are a large family of post-transcriptional regulators, which are 21-24 nt in length and play a role in a wide variety of biological processes in eukaryotes. The past few years have seen rapid progress in our understanding of miRNA biogenesis and the mechanism of action, which commonly entails a combination of target degradation and translational repression. The target degradation mediated by Argonaute-catalyzed endonucleolytic cleavage exerts a significant repressive effect on target mRNA expression, particularly during rapid developmental transitions. This review outlines the current understanding of the mechanistic aspects of this important process and discusses several different experimental approaches to identify miRNA cleavage targets.
Figures
References
-
- Krol J., Loedige I., Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. (2010);11:597–610. - PubMed
-
- Shin C. Cleavage of the star strand facilitates assembly of some microRNAs into Ago2-containing silencing complexes in mammals. Mol. Cells. (2008);26:308–313. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
