

Distinct tau prion strains propagate in cells and mice and define different tauopathies
- PMID: 24857020
- PMCID: PMC4171396
- DOI: 10.1016/j.neuron.2014.04.047
Distinct tau prion strains propagate in cells and mice and define different tauopathies
Abstract
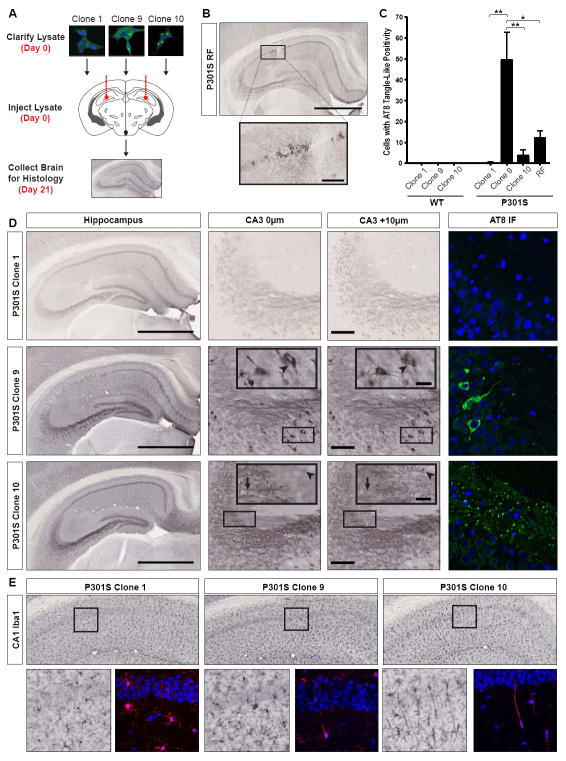
Prion-like propagation of tau aggregation might underlie the stereotyped progression of neurodegenerative tauopathies. True prions stably maintain unique conformations ("strains") in vivo that link structure to patterns of pathology. We now find that tau meets this criterion. Stably expressed tau repeat domain indefinitely propagates distinct amyloid conformations in a clonal fashion in culture. Reintroduction of tau from these lines into naive cells reestablishes identical clones. We produced two strains in vitro that induce distinct pathologies in vivo as determined by successive inoculations into three generations of transgenic mice. Immunopurified tau from these mice recreates the original strains in culture. We used the cell system to isolate tau strains from 29 patients with 5 different tauopathies, finding that different diseases are associated with different sets of strains. Tau thus demonstrates essential characteristics of a prion. This might explain the phenotypic diversity of tauopathies and could enable more effective diagnosis and therapy.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures







Comment in
-
Tau propagation, different tau phenotypes, and prion-like properties of tau.Neuron. 2014 Jun 18;82(6):1189-90. doi: 10.1016/j.neuron.2014.06.004. Neuron. 2014. PMID: 24945760
References
-
- Aguzzi A, Rajendran L. The transcellular spread of cytosolic amyloids, prions, and prionoids. Neuron. 2009;64:783–790. - PubMed
-
- Andersen P. The Hippocampus Book. Oxford University Press; 2007.
-
- Aoyagi H, Hasegawa M, Tamaoka A. Fibrillogenic nuclei composed of P301L mutant tau induce elongation of P301L tau but not wild-type tau. J Biol Chem. 2007;282:20309–20318. - PubMed
-
- Barghorn S, Zheng-Fischhöfer Q, Ackmann M, Biernat J, Bergen von M, Mandelkow EM, Mandelkow E. Structure, microtubule interactions, and paired helical filament aggregation by tau mutants of frontotemporal dementias. Biochemistry. 2000;39:11714–11721. - PubMed
-
- Bergen von M, Barghorn S, Li L, Marx A, Biernat J, Mandelkow EM, Mandelkow E. Mutations of tau protein in frontotemporal dementia promote aggregation of paired helical filaments by enhancing local beta-structure. J Biol Chem. 2001;276:48165–48174. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 1R01NS078398/NS/NINDS NIH HHS/United States
- R01 NS071835/NS/NINDS NIH HHS/United States
- 1R01NS071835/NS/NINDS NIH HHS/United States
- R01 NS078398/NS/NINDS NIH HHS/United States
- P01AG019724/AG/NIA NIH HHS/United States
- P30 NS057105/NS/NINDS NIH HHS/United States
- P50 AG023501/AG/NIA NIH HHS/United States
- 1F31NS086251/NS/NINDS NIH HHS/United States
- F31 NS086251/NS/NINDS NIH HHS/United States
- T32 GM008151/GM/NIGMS NIH HHS/United States
- MR/K022105/1/MRC_/Medical Research Council/United Kingdom
- P50AG023501/AG/NIA NIH HHS/United States
- P01 AG019724/AG/NIA NIH HHS/United States
- P30NS057105/NS/NINDS NIH HHS/United States
- F30 AG048653/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
