

Role of lipid rafts and GM1 in the segregation and processing of prion protein
- PMID: 24859148
- PMCID: PMC4032283
- DOI: 10.1371/journal.pone.0098344
Role of lipid rafts and GM1 in the segregation and processing of prion protein
Abstract
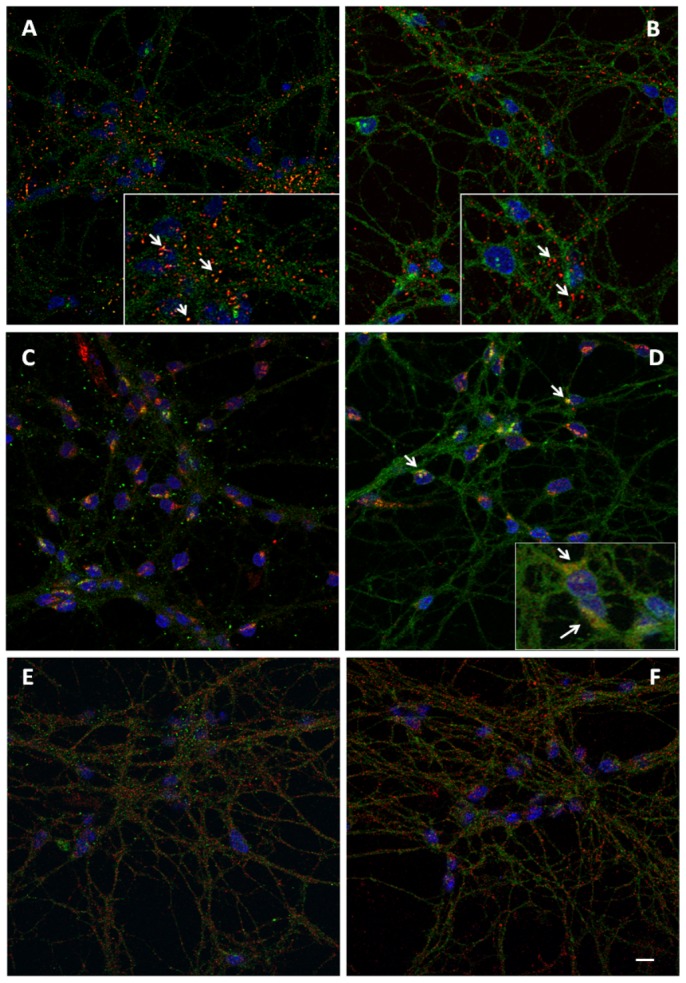
The prion protein (PrPC) is highly expressed within the nervous system. Similar to other GPI-anchored proteins, PrPC is found in lipid rafts, membrane domains enriched in cholesterol and sphingolipids. PrPC raft association, together with raft lipid composition, appears essential for the conversion of PrPC into the scrapie isoform PrPSc, and the development of prion disease. Controversial findings were reported on the nature of PrPC-containing rafts, as well as on the distribution of PrPC between rafts and non-raft membranes. We investigated PrPC/ganglioside relationships and their influence on PrPC localization in a neuronal cellular model, cerebellar granule cells. Our findings argue that in these cells at least two PrPC conformations coexist: in lipid rafts PrPC is present in the native folding (α-helical), stabilized by chemico-physical condition, while it is mainly present in other membrane compartments in a PrPSc-like conformation. We verified, by means of antibody reactivity and circular dichroism spectroscopy, that changes in lipid raft-ganglioside content alters PrPC conformation and interaction with lipid bilayers, without modifying PrPC distribution or cleavage. Our data provide new insights into the cellular mechanism of prion conversion and suggest that GM1-prion protein interaction at the cell surface could play a significant role in the mechanism predisposing to pathology.
Conflict of interest statement
Figures








Similar articles
-
Binding of prion proteins to lipid membranes.Biochem Biophys Res Commun. 2004 Jan 16;313(3):559-67. doi: 10.1016/j.bbrc.2003.12.004. Biochem Biophys Res Commun. 2004. PMID: 14697227
-
The cellular prion protein with a monoacylated glycosylphosphatidylinositol anchor modifies cell membranes, inhibits cell signaling and reduces prion formation.Prion. 2011 Apr-Jun;5(2):65-8. doi: 10.4161/pri.5.2.16095. Epub 2011 Apr 1. Prion. 2011. PMID: 21738009 Free PMC article.
-
Glypican-1 mediates both prion protein lipid raft association and disease isoform formation.PLoS Pathog. 2009 Nov;5(11):e1000666. doi: 10.1371/journal.ppat.1000666. Epub 2009 Nov 20. PLoS Pathog. 2009. PMID: 19936054 Free PMC article.
-
The role of rafts in the fibrillization and aggregation of prions.Chem Phys Lipids. 2006 Jun;141(1-2):66-71. doi: 10.1016/j.chemphyslip.2006.02.022. Epub 2006 Mar 27. Chem Phys Lipids. 2006. PMID: 16647049 Review.
-
The prion protein and lipid rafts.Mol Membr Biol. 2006 Jan-Feb;23(1):89-99. doi: 10.1080/09687860500449994. Mol Membr Biol. 2006. PMID: 16611584 Review.
Cited by
-
Mechanism of scrapie prion precipitation with phosphotungstate anions.ACS Chem Biol. 2015 May 15;10(5):1269-77. doi: 10.1021/cb5006239. Epub 2015 Feb 19. ACS Chem Biol. 2015. PMID: 25695325 Free PMC article.
-
Changes in Membrane Protein Clustering in Peripheral Lymphocytes in an Animal Model of Depression Parallel Those Observed in Naïve Depression Patients: Implications for the Development of Novel Biomarkers of Depression.Front Pharmacol. 2018 Oct 15;9:1149. doi: 10.3389/fphar.2018.01149. eCollection 2018. Front Pharmacol. 2018. PMID: 30374301 Free PMC article.
-
Structural and dynamic views of GM1 ganglioside.Glycoconj J. 2015 May;32(3-4):105-12. doi: 10.1007/s10719-015-9587-5. Epub 2015 May 1. Glycoconj J. 2015. PMID: 25931034 Review.
-
PrP C as a Transducer of Physiological and Pathological Signals.Front Mol Neurosci. 2021 Nov 22;14:762918. doi: 10.3389/fnmol.2021.762918. eCollection 2021. Front Mol Neurosci. 2021. PMID: 34880726 Free PMC article. Review.
-
Prion Protein in Stem Cells: A Lipid Raft Component Involved in the Cellular Differentiation Process.Int J Mol Sci. 2020 Jun 11;21(11):4168. doi: 10.3390/ijms21114168. Int J Mol Sci. 2020. PMID: 32545192 Free PMC article. Review.
References
-
- Oesch B, Westaway D, Wälchli M, McKinley MP, Kent SBH, et al. (1985) A cellular gene encodes scrapie PrP 27–30 protein. Cell 40: 735–746 doi:10.1016/0092-8674(85)90333-2 - DOI - PubMed
-
- Biasini E, Turnbaugh JA, Unterberger U, Harris DA (2012) Prion protein at the crossroads of physiology and disease. Trends Neurosci 36: 92–103 doi: 10.1016/j.tins.2011.10.002 - DOI - PMC - PubMed
-
- Simons K, Ikonen E (1997) Functional rafts in cell membranes. Nature 387: 569–572 doi:10.1038/42408 - DOI - PubMed
-
- Allen JA, Halverson-Tamboli RA, Rasenick MM (2007) Lipid raft microdomains and neurotransmitter signalling. Nature, Neuroscience 8: 128–140 doi:10.1038/nrn2059 - DOI - PubMed
-
- Prusiner SB (1998) Prions. Proc Natl Acad Sci USA. 95: 13363–13383 doi: 10.1073/pnas.95.23.13363 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
