

Protective effect of astaxanthin on liver fibrosis through modulation of TGF-β1 expression and autophagy
- PMID: 24860243
- PMCID: PMC4016904
- DOI: 10.1155/2014/954502
Protective effect of astaxanthin on liver fibrosis through modulation of TGF-β1 expression and autophagy
Abstract
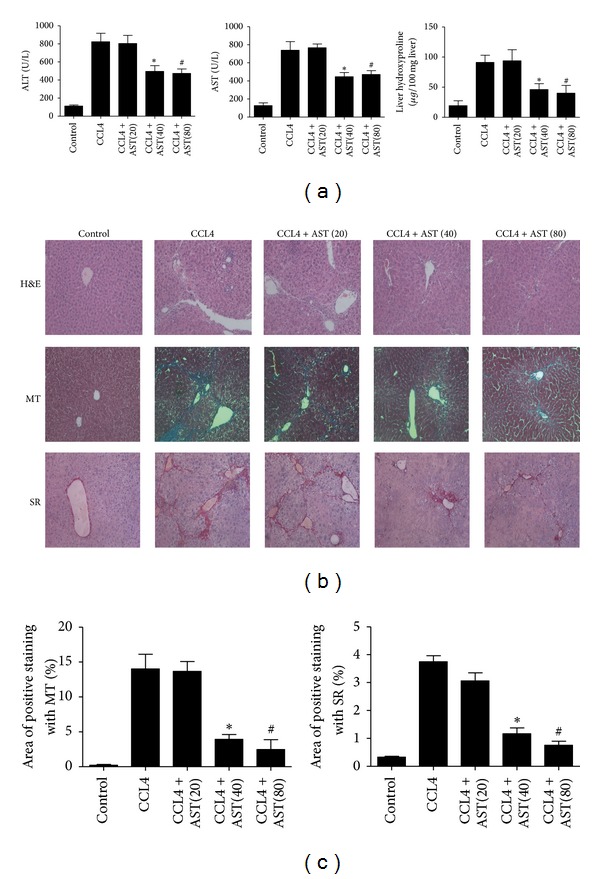
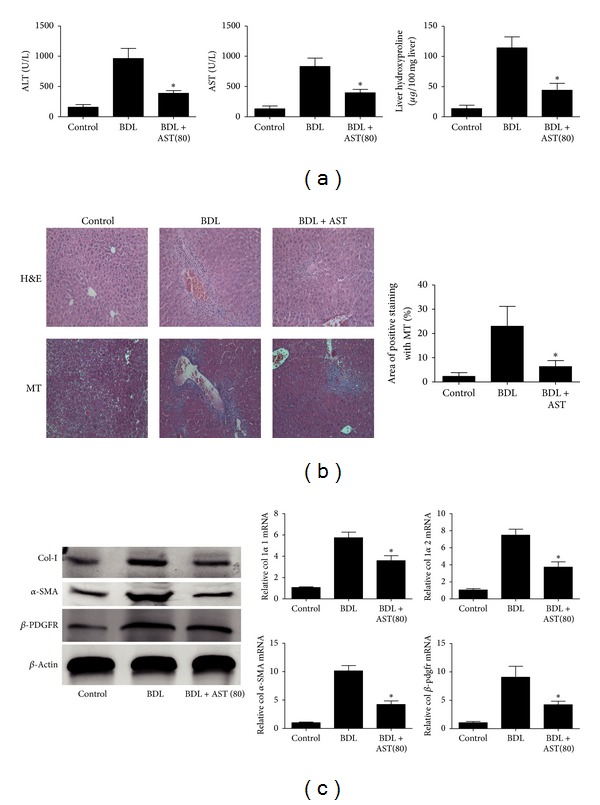
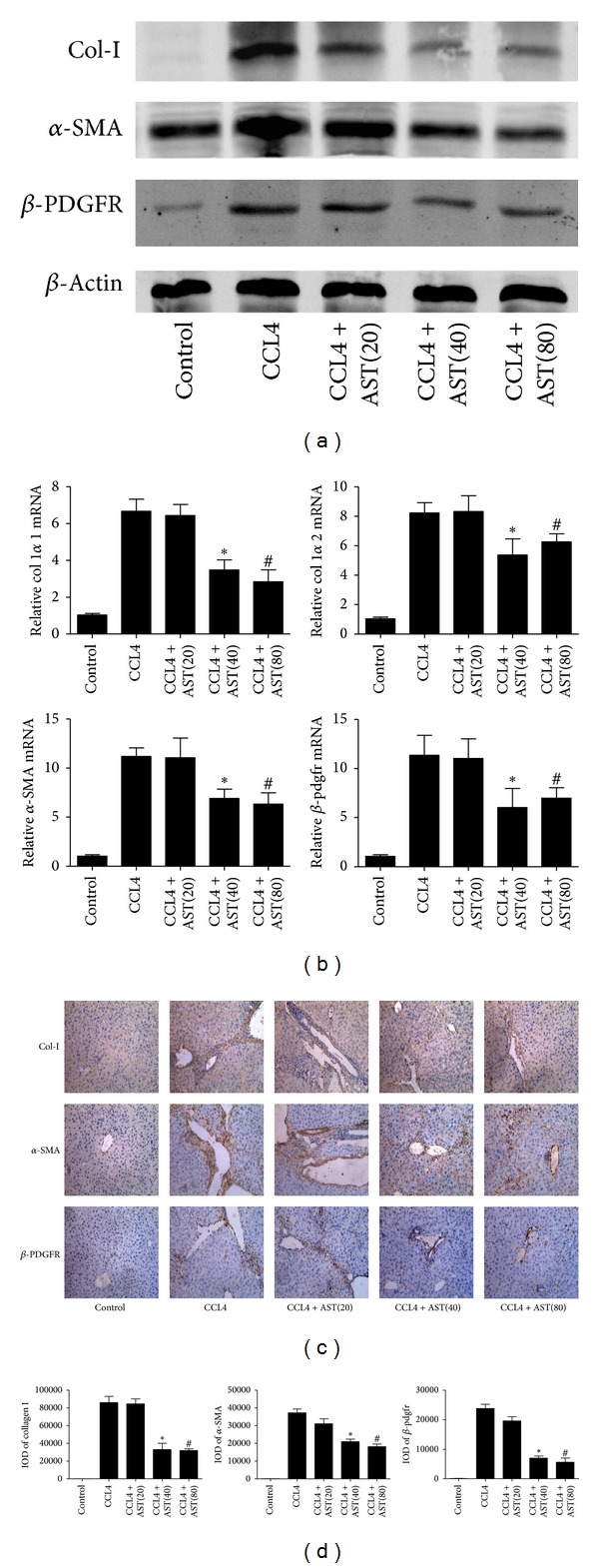
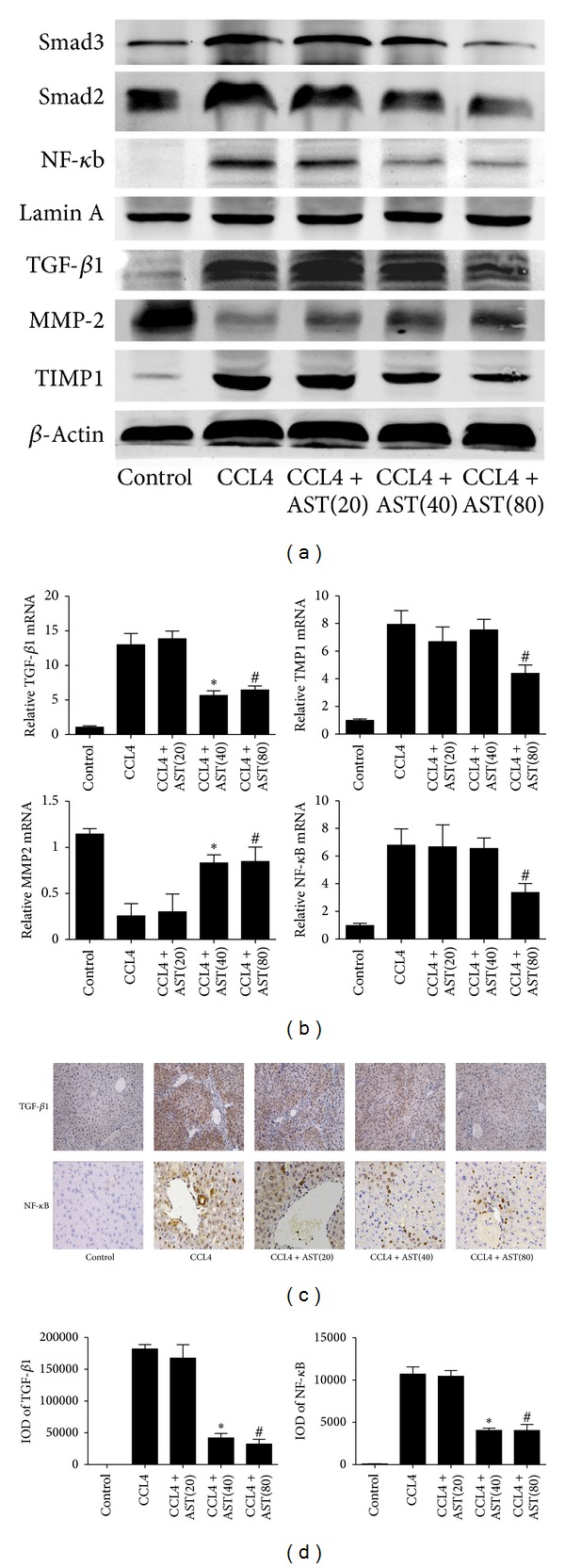
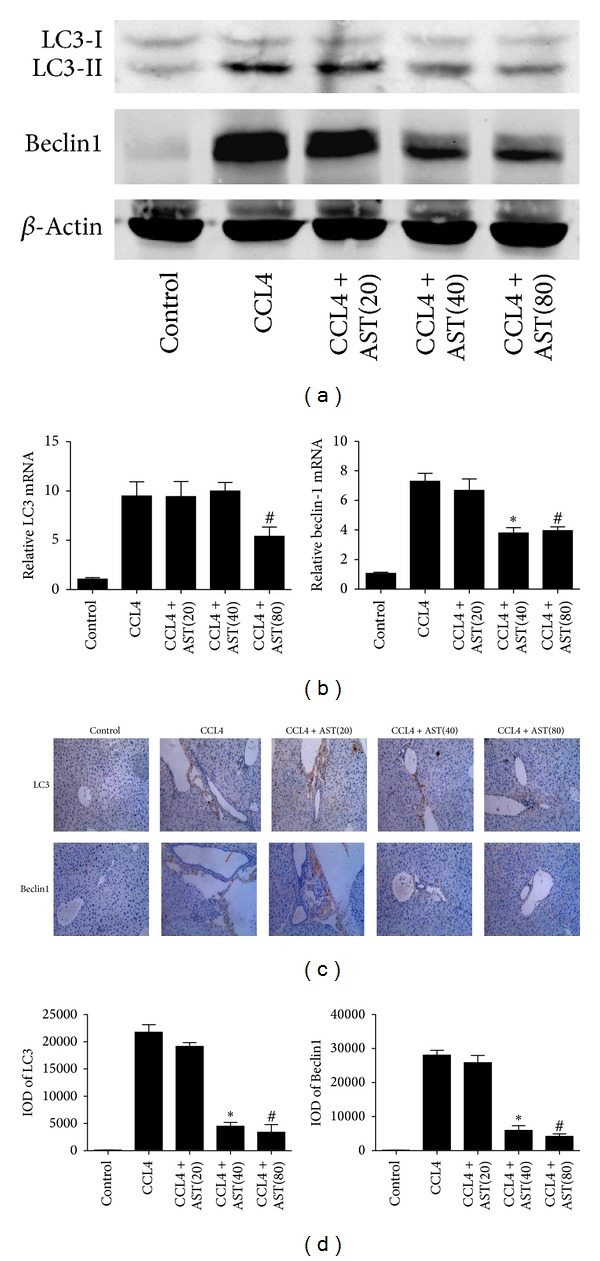
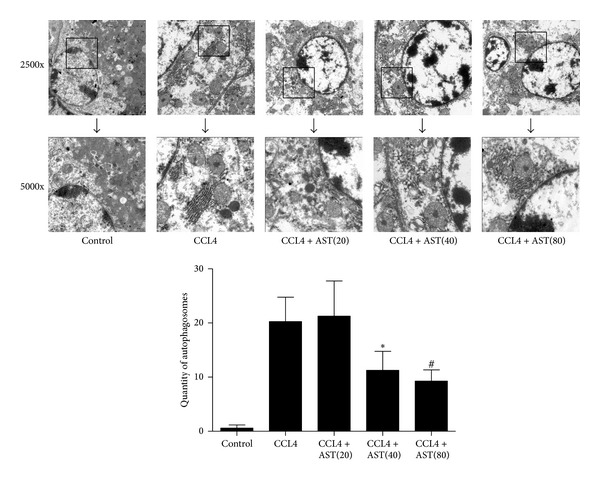
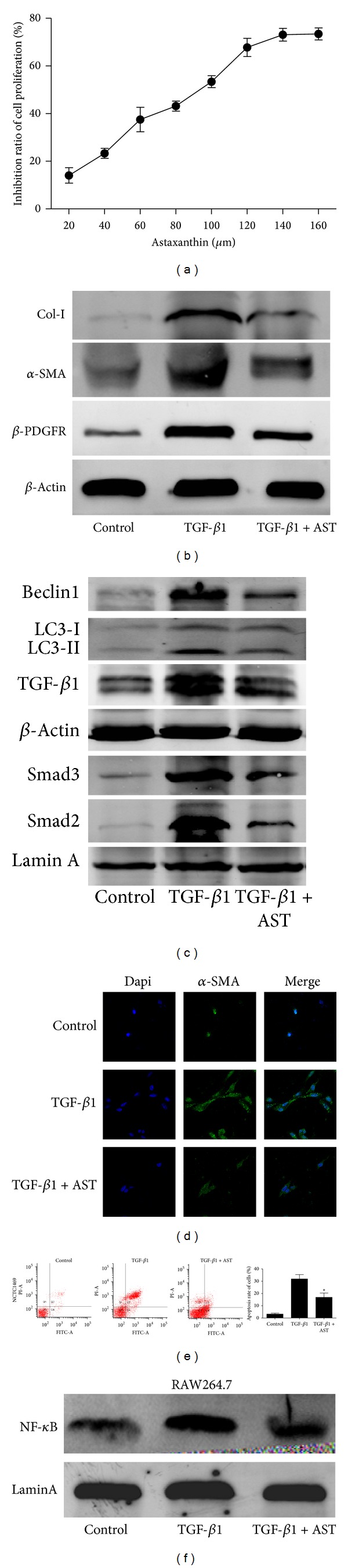
Liver fibrosis is a common pathway leading to cirrhosis and a worldwide clinical issue. Astaxanthin is a red carotenoid pigment with antioxidant, anticancer, and anti-inflammatory properties. The aim of this study was to investigate the effect of astaxanthin on liver fibrosis and its potential protective mechanisms. Liver fibrosis was induced in a mouse model using CCL4 (intraperitoneal injection, three times a week for 8 weeks), and astaxanthin was administered everyday at three doses (20, 40, and 80 mg/kg). Pathological results indicated that astaxanthin significantly improved the pathological lesions of liver fibrosis. The levels of alanine aminotransferase aspartate aminotransferase and hydroxyproline were also significantly decreased by astaxanthin. The same results were confirmed in bile duct liagtion, (BDL) model. In addition, astaxanthin inhibited hepatic stellate cells (HSCs) activation and formation of extracellular matrix (ECM) by decreasing the expression of NF-κB and TGF-β1 and maintaining the balance between MMP2 and TIMP1. In addition, astaxanthin reduced energy production in HSCs by downregulating the level of autophagy. These results were simultaneously confirmed in vivo and in vitro. In conclusion, our study showed that 80 mg/kg astaxanthin had a significant protective effect on liver fibrosis by suppressing multiple profibrogenic factors.
Figures
References
-
- Kisseleva T, Brenner DA. Role of hepatic stellate cells in fibrogenesis and the reversal of fibrosis. Journal of Gastroenterology and Hepatology. 2007;22(1):S73–S78. - PubMed
-
- Tacke F, Weiskirchen R. Update on hepatic stellate cells: pathogenic role in liver fibrosis and novel isolation techniques. Expert Review of Gastroenterology and Hepatology. 2012;6(1):67–80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
