

Intrinsic properties and neuropharmacology of midline paraventricular thalamic nucleus neurons
- PMID: 24860449
- PMCID: PMC4029024
- DOI: 10.3389/fnbeh.2014.00132
Intrinsic properties and neuropharmacology of midline paraventricular thalamic nucleus neurons
Abstract
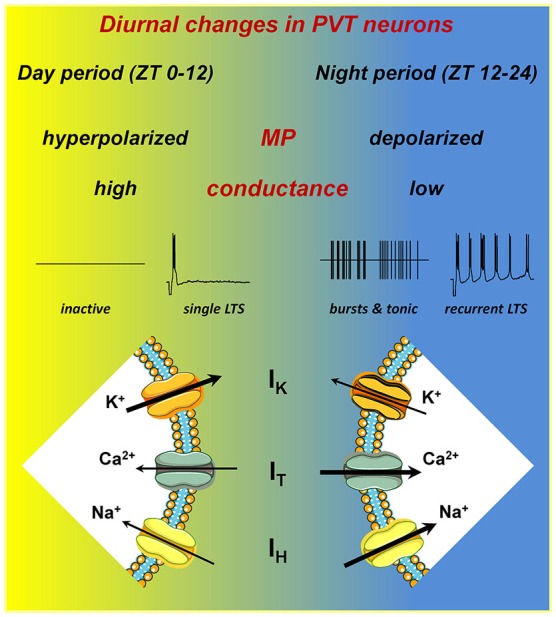
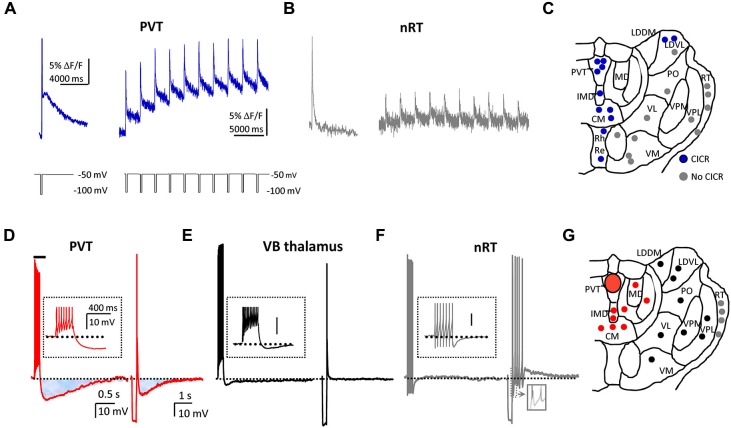
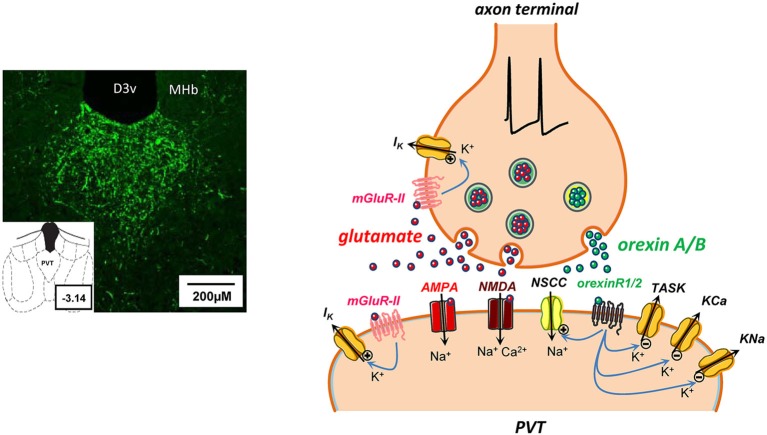
Neurons in the midline and intralaminar thalamic nuclei are components of an interconnected brainstem, limbic and prefrontal cortex neural network that is engaged during arousal, vigilance, motivated and addictive behaviors, and stress. To better understand the cellular mechanisms underlying these functions, here we review some of the recently characterized electrophysiological and neuropharmacological properties of neurons in the paraventricular thalamic nucleus (PVT), derived from whole cell patch clamp recordings in acute rat brain slice preparations. PVT neurons display firing patterns and ionic conductances (IT and IH) that exhibit significant diurnal change. Their resting membrane potential (RMP) is maintained by various ionic conductances that include inward rectifier (Kir), hyperpolarization-activated nonselective cation (HCN) and TWIK-related acid sensitive (TASK) K(+) channels. Firing patterns are regulated by high voltage-activated (HVA) and low voltage-activated (LVA) Ca(2+) conductances. Moreover, transient receptor potential (TRP)-like nonselective cation channels together with Ca(2+)- and Na(+)-activated K(+) conductances (KCa; KNa) contribute to unique slow afterhyperpolarizing potentials (sAHPs) that are generally not detectable in lateral thalamic or reticular thalamic nucleus neurons. The excitability of PVT neurons is also modulated by activation of neurotransmitter receptors associated with afferent pathways to PVT and other thalamic midline nuclei. We report on receptor-mediated actions of GABA, glutamate, monoamines and several neuropeptides: arginine vasopressin, gastrin-releasing peptide, thyrotropin releasing hormone and the orexins (hypocretins). This review represents an initial survey of intrinsic and transmitter-sensitive ionic conductances that are deemed to be unique to this population of midline thalamic neurons, information that is fundamental to an appreciation of the role these thalamic neurons may play in normal central nervous system (CNS) physiology and in CNS disorders that involve the dorsomedial thalamus.
Keywords: burst firing; diurnal and seasonal changes; electrophysiology; midline thalamic nuclei; peptides.
Figures
Similar articles
-
Gastrin-releasing peptide acts via postsynaptic BB2 receptors to modulate inward rectifier K+ and TRPV1-like conductances in rat paraventricular thalamic neurons.J Physiol. 2013 Apr 1;591(7):1823-39. doi: 10.1113/jphysiol.2012.249227. Epub 2013 Jan 28. J Physiol. 2013. PMID: 23359674 Free PMC article.
-
Novel coupling between TRPC-like and KNa channels modulates low threshold spike-induced afterpotentials in rat thalamic midline neurons.Neuropharmacology. 2014 Nov;86:88-96. doi: 10.1016/j.neuropharm.2014.06.023. Epub 2014 Jul 9. Neuropharmacology. 2014. PMID: 25014020
-
Calcium influx through N-type channels and activation of SK and TRP-like channels regulates tonic firing of neurons in rat paraventricular thalamus.J Neurophysiol. 2013 Nov;110(10):2450-64. doi: 10.1152/jn.00363.2013. Epub 2013 Sep 4. J Neurophysiol. 2013. PMID: 24004531
-
Limbic thalamus and state-dependent behavior: The paraventricular nucleus of the thalamic midline as a node in circadian timing and sleep/wake-regulatory networks.Neurosci Biobehav Rev. 2015 Jul;54:3-17. doi: 10.1016/j.neubiorev.2014.11.021. Epub 2014 Dec 3. Neurosci Biobehav Rev. 2015. PMID: 25479103 Review.
-
The intralaminar and midline nuclei of the thalamus. Anatomical and functional evidence for participation in processes of arousal and awareness.Brain Res Brain Res Rev. 2002 Sep;39(2-3):107-40. doi: 10.1016/s0165-0173(02)00181-9. Brain Res Brain Res Rev. 2002. PMID: 12423763 Review.
Cited by
-
The paraventricular nucleus of the thalamus: an integrative node underlying homeostatic behavior.Trends Neurosci. 2021 Jul;44(7):538-549. doi: 10.1016/j.tins.2021.03.001. Epub 2021 Mar 25. Trends Neurosci. 2021. PMID: 33775435 Free PMC article. Review.
-
Sex differences in electrophysiological properties and voltage-gated ion channel expression in the paraventricular thalamic nucleus following repeated stress.Biol Sex Differ. 2022 Sep 27;13(1):51. doi: 10.1186/s13293-022-00460-0. Biol Sex Differ. 2022. PMID: 36163074 Free PMC article.
-
A Neural Circuit from Thalamic Paraventricular Nucleus to Central Amygdala for the Facilitation of Neuropathic Pain.J Neurosci. 2020 Oct 7;40(41):7837-7854. doi: 10.1523/JNEUROSCI.2487-19.2020. Epub 2020 Sep 21. J Neurosci. 2020. PMID: 32958568 Free PMC article.
-
Circuits and components of delta wave regulation.Brain Res Bull. 2022 Oct 1;188:223-232. doi: 10.1016/j.brainresbull.2022.06.006. Epub 2022 Jun 20. Brain Res Bull. 2022. PMID: 35738502 Free PMC article. Review.
-
Research progress of the paraventricular thalamus in the regulation of sleep-wake and emotional behaviors.Ibrain. 2022 Apr 30;8(2):219-226. doi: 10.1002/ibra.12034. eCollection 2022 Summer. Ibrain. 2022. PMID: 37786895 Free PMC article. Review.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous