

Competition as a way of life for H(+)-coupled antiporters
- PMID: 24862284
- PMCID: PMC4072998
- DOI: 10.1016/j.jmb.2014.05.020
Competition as a way of life for H(+)-coupled antiporters
Abstract
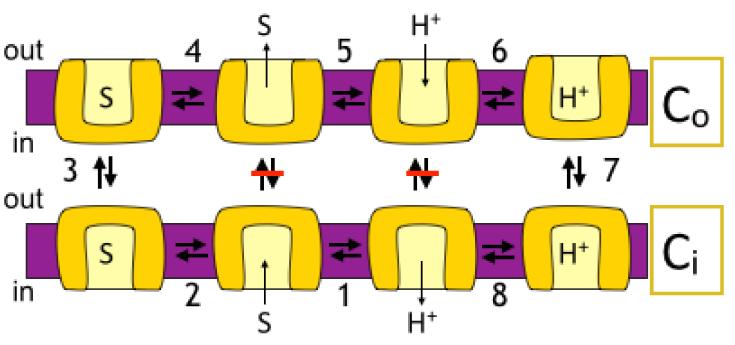
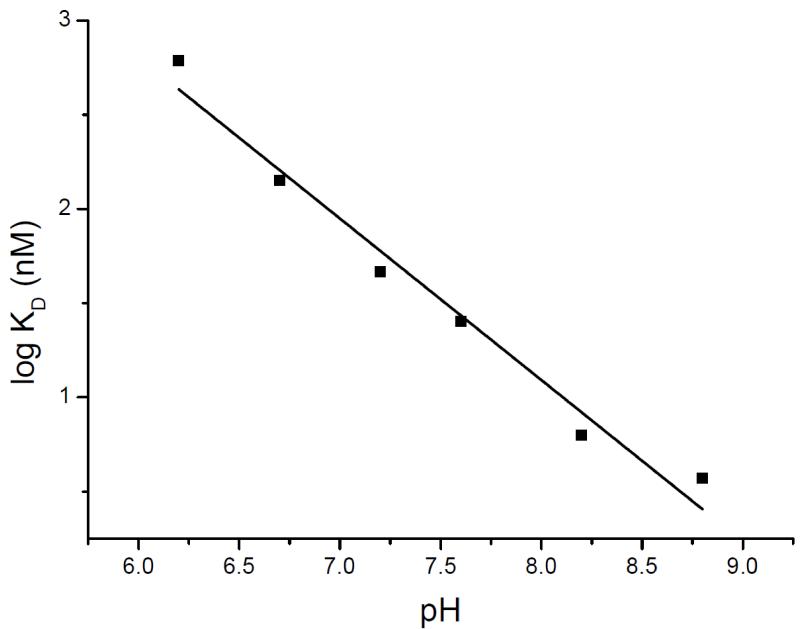
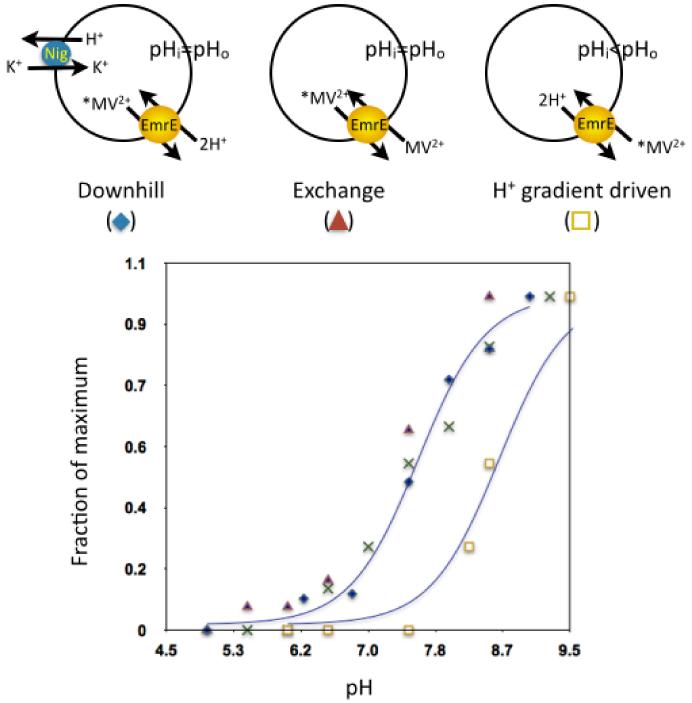
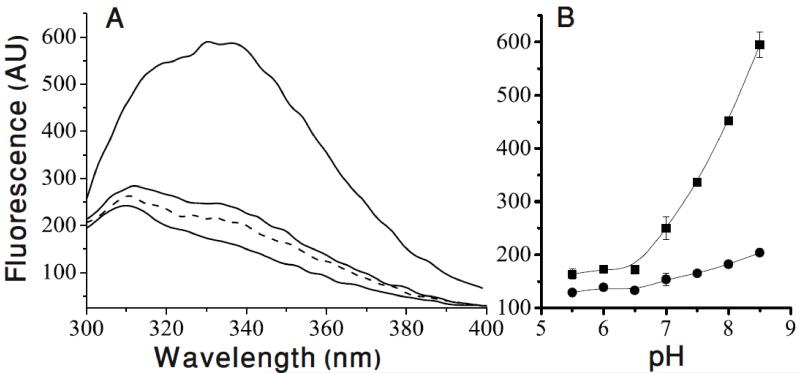
Antiporters are ubiquitous membrane proteins that catalyze obligatory exchange between two or more substrates across a membrane in opposite directions. Some utilize proton electrochemical gradients generated by primary pumps by coupling the downhill movement of one or more protons to the movement of a substrate. Since the direction of the proton gradient usually favors proton movement toward the cytoplasm, their function results in removal of substrates other than protons from the cytoplasm, either into acidic intracellular compartments or out to the medium. H(+)-coupled antiporters play central roles in living organisms, for example, storage of neurotransmitter and other small molecules, resistance to antibiotics, homeostasis of ionic content and more. Biochemical and structural data support a general mechanism for H(+)-coupled antiporters whereby the substrate and the protons cannot bind simultaneously to the protein. In several cases, it was shown that the binding sites overlap, and therefore, there is a direct competition between the protons and the substrate. In others, the "competition" seems to be indirect and it is most likely achieved by allosteric mechanisms. The pKa of one or more carboxyls in the protein must be tuned appropriately in order to ensure the feasibility of such a mechanism. In this review, I discuss in detail the case of EmrE, a multidrug transporter from Escherichia coli and evaluate the information available for other H(+)-coupled antiporters.
Keywords: coupling mechanism; membrane proteins; multidrug transporters; transport; vesicular neurotransmitter transporters.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Fluman N, Bibi E. Bacterial multidrug transport through the lens of the major facilitator superfamily. Biochim Biophys Acta. 2009;1794:738–47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
