

Design of synthetic yeast promoters via tuning of nucleosome architecture
- PMID: 24862902
- PMCID: PMC4064463
- DOI: 10.1038/ncomms5002
Design of synthetic yeast promoters via tuning of nucleosome architecture
Abstract
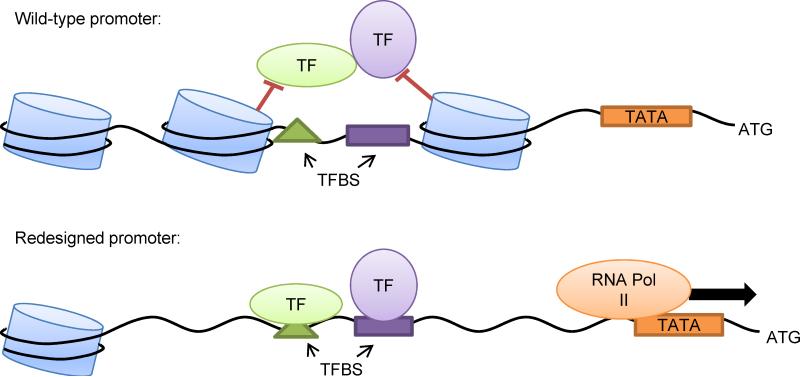
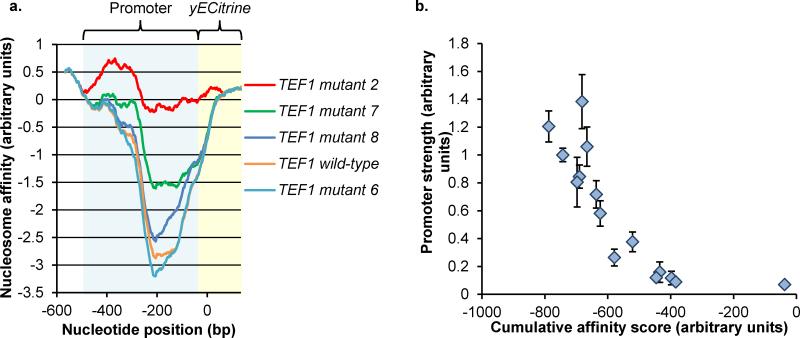
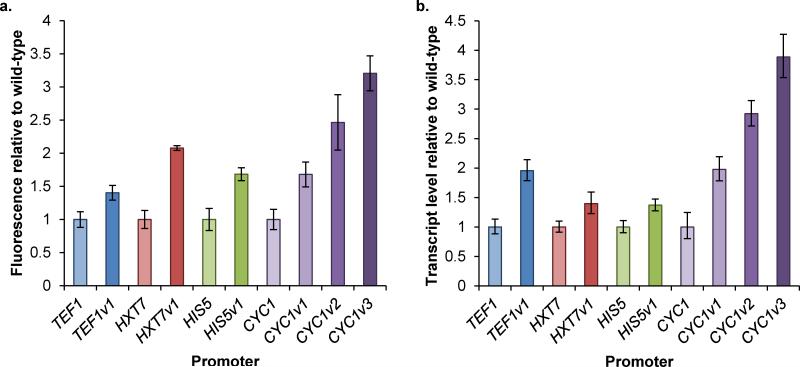
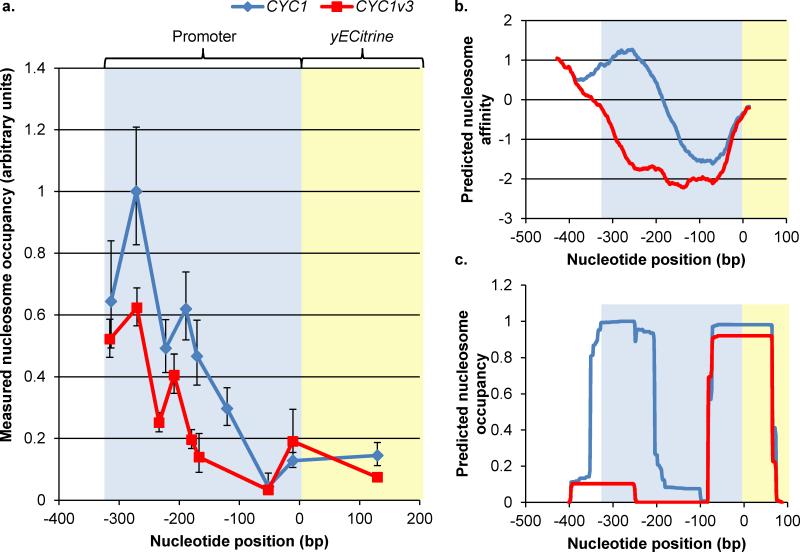
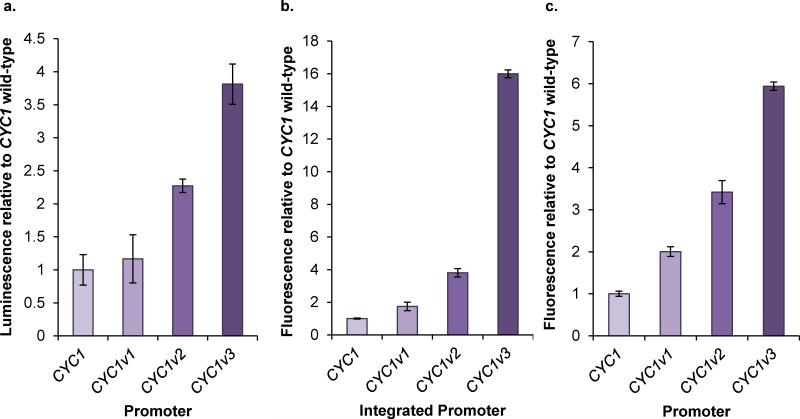
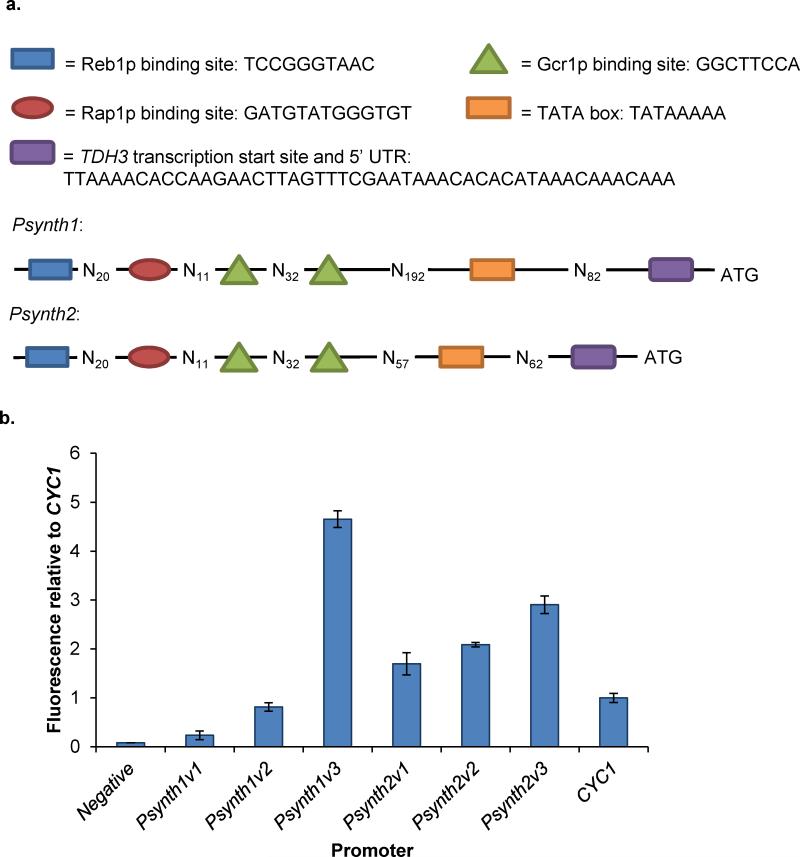
Model-based design of biological parts is a critical goal of synthetic biology, especially for eukaryotes. Here we demonstrate that nucleosome architecture can have a role in defining yeast promoter activity and utilize a computationally-guided approach that can enable both the redesign of endogenous promoter sequences and the de novo design of synthetic promoters. Initially, we use our approach to reprogram native promoters for increased expression and evaluate their performance in various genetic contexts. Increases in expression ranging from 1.5- to nearly 6-fold in a plasmid-based system and up to 16-fold in a genomic context were obtained. Next, we demonstrate that, in a single design cycle, it is possible to create functional, purely synthetic yeast promoters that achieve substantial expression levels (within the top sixth percentile among native yeast promoters). In doing so, this work establishes a unique DNA-level specification of promoter activity and demonstrates predictive design of synthetic parts.
Figures

References
-
- Crook N, Alper HS. Model-based design of synthetic, biological systems. Chemical Engineering Science. 2013;103:2–11.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
