

Counter-regulation of T cell effector function by differentially activated p38
- PMID: 24863062
- PMCID: PMC4042639
- DOI: 10.1084/jem.20131917
Counter-regulation of T cell effector function by differentially activated p38
Abstract
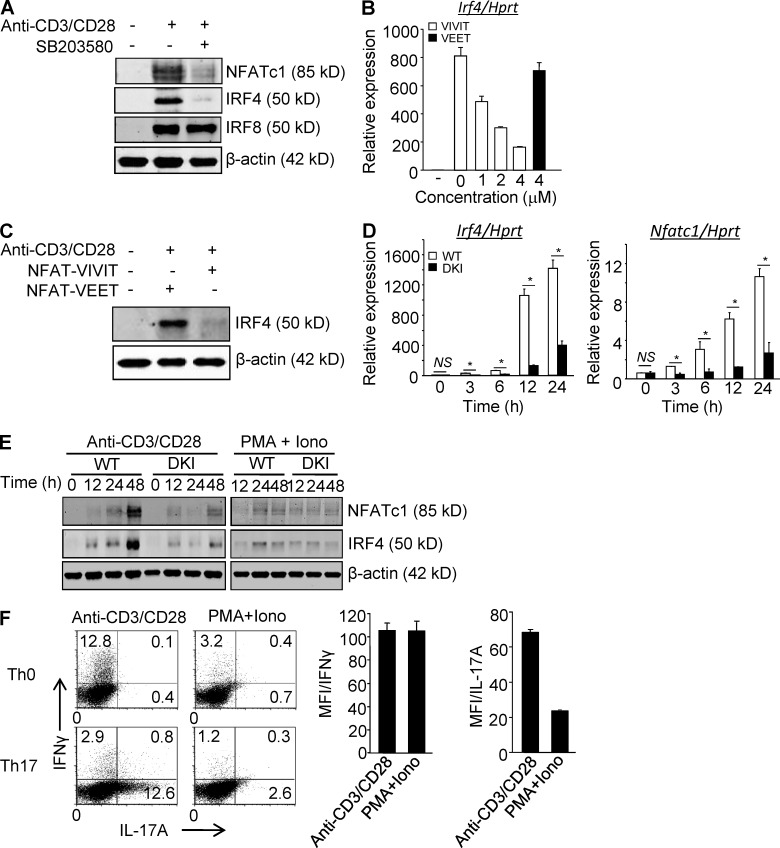
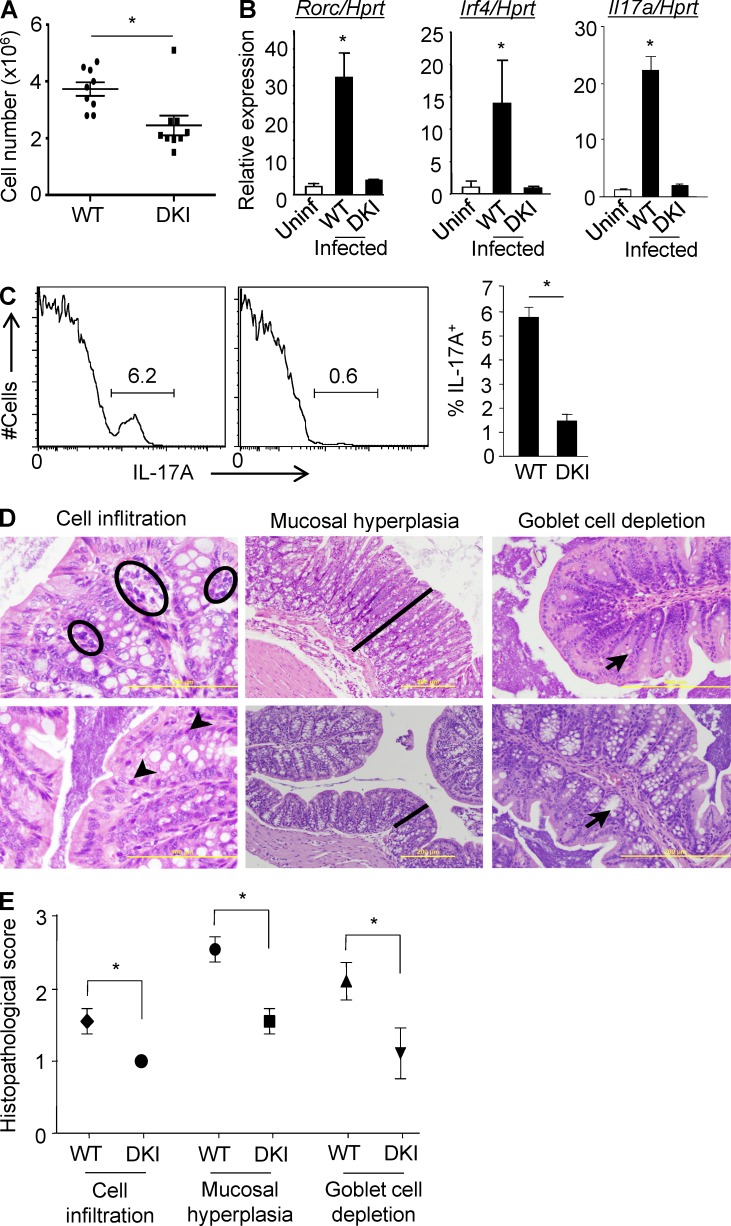
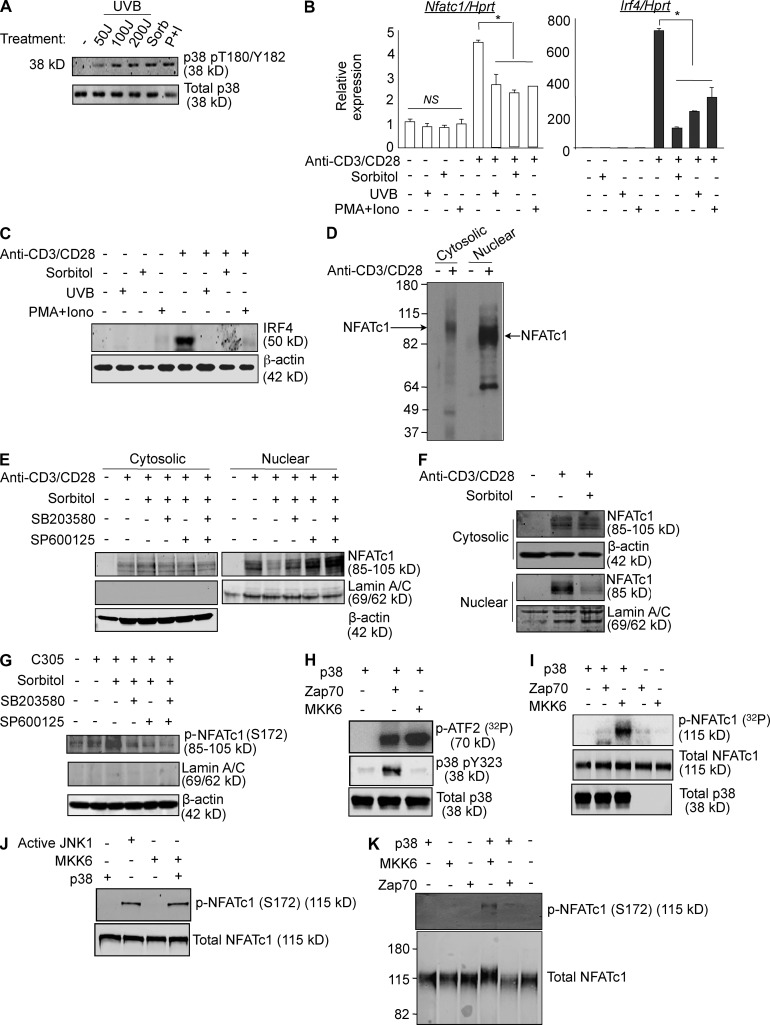
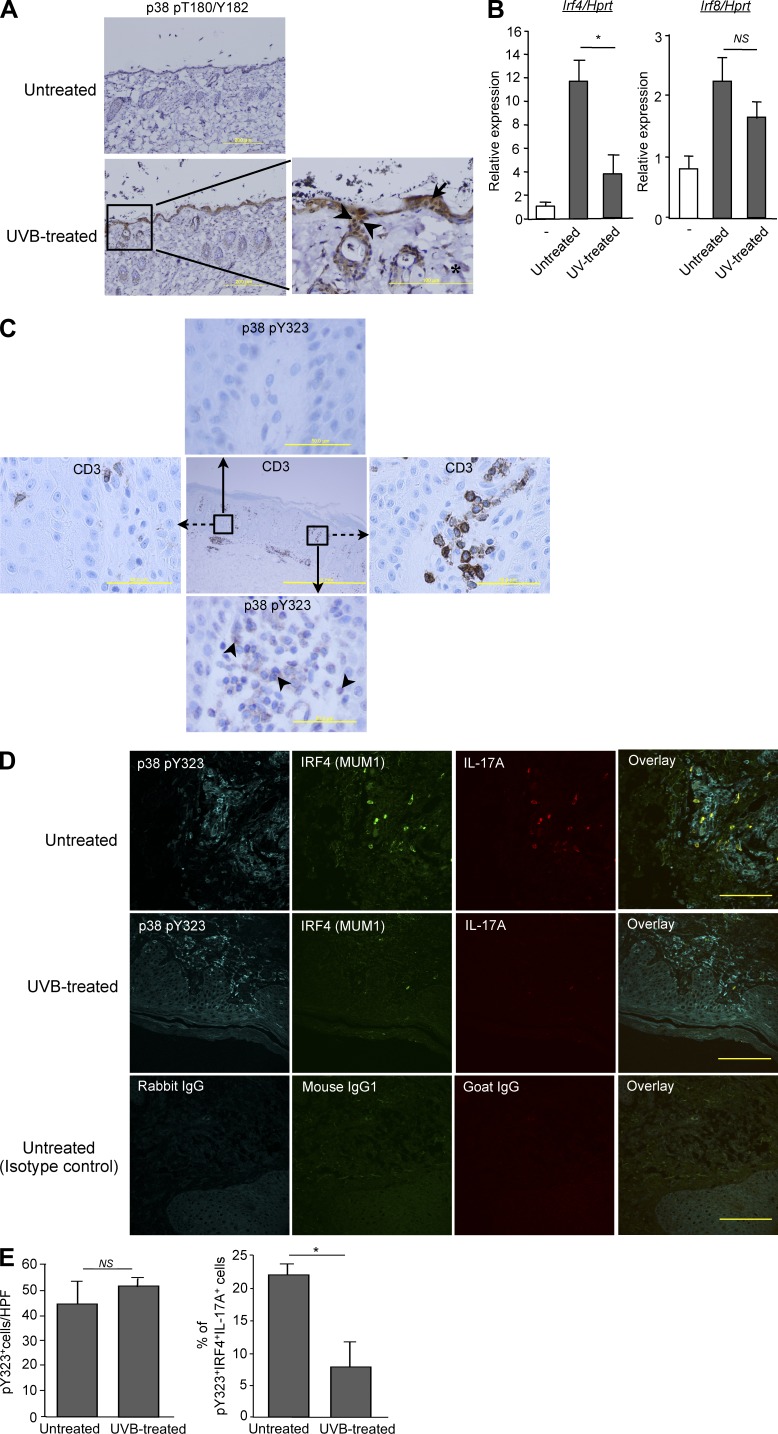
Unlike the MAP kinase (MAPK) cascade that phosphorylates p38 on the activation loop, T cell receptor (TCR) signaling results in phosphorylation on Tyr-323 (pY323, alternative pathway). Using mice expressing p38α and p38β with Y323F substitutions, we show that alternatively but not MAPK cascade-activated p38 up-regulates the transcription factors NFATc1 and IRF4, which are required for proliferation and cytokine production. Conversely, activation of p38 with UV or osmotic shock mitigated TCR-mediated activation by phosphorylation and cytoplasmic retention of NFATc1. Notably, UVB treatment of human psoriatic lesions reduced skin-infiltrating p38 pY323(+) T cell IRF4 and IL-17 production. Thus, distinct mechanisms of p38 activation converge on NFATc1 with opposing effects on T cell immunity, which may underlie the beneficial effect of phototherapy on psoriasis.
Figures

Similar articles
-
Lack of the T cell-specific alternative p38 activation pathway reduces autoimmunity and inflammation.Blood. 2011 Sep 22;118(12):3280-9. doi: 10.1182/blood-2011-01-333039. Epub 2011 Jun 28. Blood. 2011. PMID: 21715315 Free PMC article.
-
Genetic disruption of p38alpha Tyr323 phosphorylation prevents T-cell receptor-mediated p38alpha activation and impairs interferon-gamma production.Blood. 2009 Mar 5;113(10):2229-37. doi: 10.1182/blood-2008-04-153304. Epub 2008 Nov 14. Blood. 2009. PMID: 19011223 Free PMC article.
-
Selective inhibition of the p38 alternative activation pathway in infiltrating T cells inhibits pancreatic cancer progression.Nat Med. 2015 Nov;21(11):1337-43. doi: 10.1038/nm.3957. Epub 2015 Oct 19. Nat Med. 2015. PMID: 26479921 Free PMC article.
-
Activating p38 MAPK: new tricks for an old kinase.Cell Cycle. 2005 Sep;4(9):1189-92. doi: 10.4161/cc.4.9.2043. Epub 2005 Sep 20. Cell Cycle. 2005. PMID: 16103752 Review.
-
The role of p38 MAPK in the aetiopathogenesis of psoriasis and psoriatic arthritis.Clin Dev Immunol. 2013;2013:569751. doi: 10.1155/2013/569751. Epub 2013 Sep 12. Clin Dev Immunol. 2013. PMID: 24151518 Free PMC article. Review.
Cited by
-
Roles of p38α mitogen-activated protein kinase in mouse models of inflammatory diseases and cancer.FEBS J. 2015 May;282(10):1841-57. doi: 10.1111/febs.13250. Epub 2015 Mar 16. FEBS J. 2015. PMID: 25728574 Free PMC article. Review.
-
Mixed lineage kinase 3 and CD70 cooperation sensitize trastuzumab-resistant HER2+ breast cancer by ceramide-loaded nanoparticles.Proc Natl Acad Sci U S A. 2022 Sep 20;119(38):e2205454119. doi: 10.1073/pnas.2205454119. Epub 2022 Sep 12. Proc Natl Acad Sci U S A. 2022. PMID: 36095190 Free PMC article.
-
cIAP1/2 inhibition synergizes with TNF inhibition in autoimmunity by down-regulating IL-17A and inducing Tregs.Sci Adv. 2019 May 1;5(5):eaaw5422. doi: 10.1126/sciadv.aaw5422. eCollection 2019 May. Sci Adv. 2019. PMID: 31049403 Free PMC article.
-
Targeting protein modification: a new direction for immunotherapy of pancreatic cancer.Int J Biol Sci. 2025 Jan 1;21(1):63-74. doi: 10.7150/ijbs.101861. eCollection 2025. Int J Biol Sci. 2025. PMID: 39744438 Free PMC article. Review.
-
Multi-Kinase Inhibitor with Anti-p38γ Activity in Cutaneous T-Cell Lymphoma.J Invest Dermatol. 2018 Nov;138(11):2377-2387. doi: 10.1016/j.jid.2018.04.030. Epub 2018 Jul 14. J Invest Dermatol. 2018. PMID: 29758280 Free PMC article.
References
-
- Aragão A.Z., Nogueira M.L., Granato D.C., Simabuco F.M., Honorato R.V., Hoffman Z., Yokoo S., Laurindo F.R., Squina F.M., Zeri A.C., et al. 2012. Identification of novel interaction between ADAM17 (a disintegrin and metalloprotease 17) and thioredoxin-1. J. Biol. Chem. 287:43071–43082 10.1074/jbc.M112.364513 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
