

Tomato PYR/PYL/RCAR abscisic acid receptors show high expression in root, differential sensitivity to the abscisic acid agonist quinabactin, and the capability to enhance plant drought resistance
- PMID: 24863435
- PMCID: PMC4112642
- DOI: 10.1093/jxb/eru219
Tomato PYR/PYL/RCAR abscisic acid receptors show high expression in root, differential sensitivity to the abscisic acid agonist quinabactin, and the capability to enhance plant drought resistance
Abstract
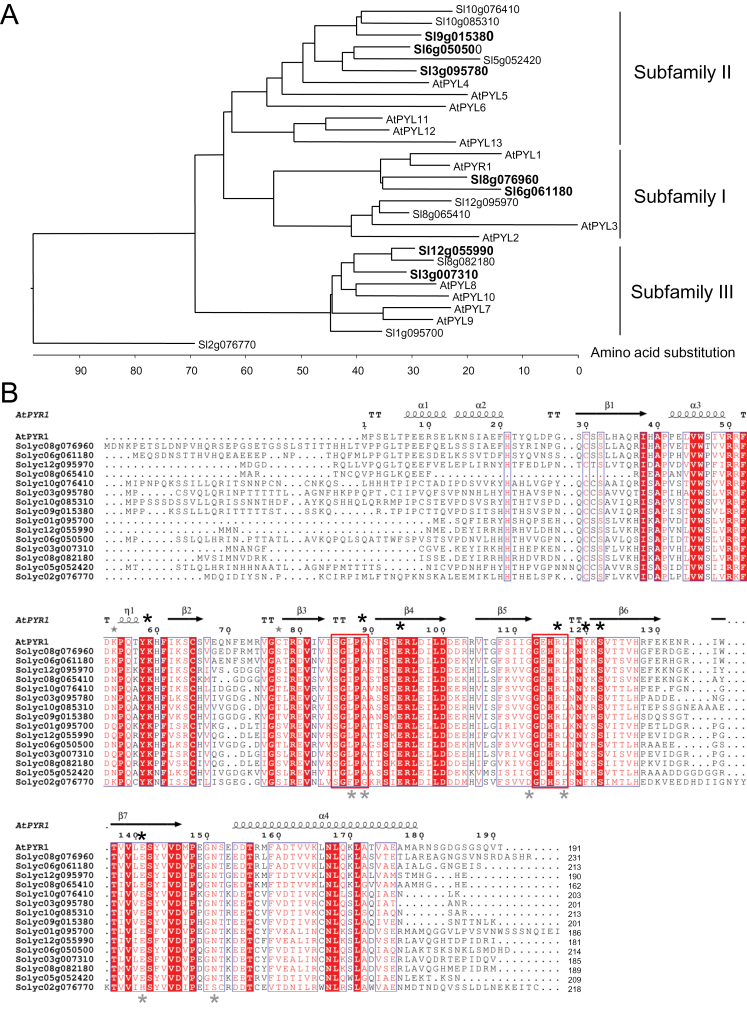
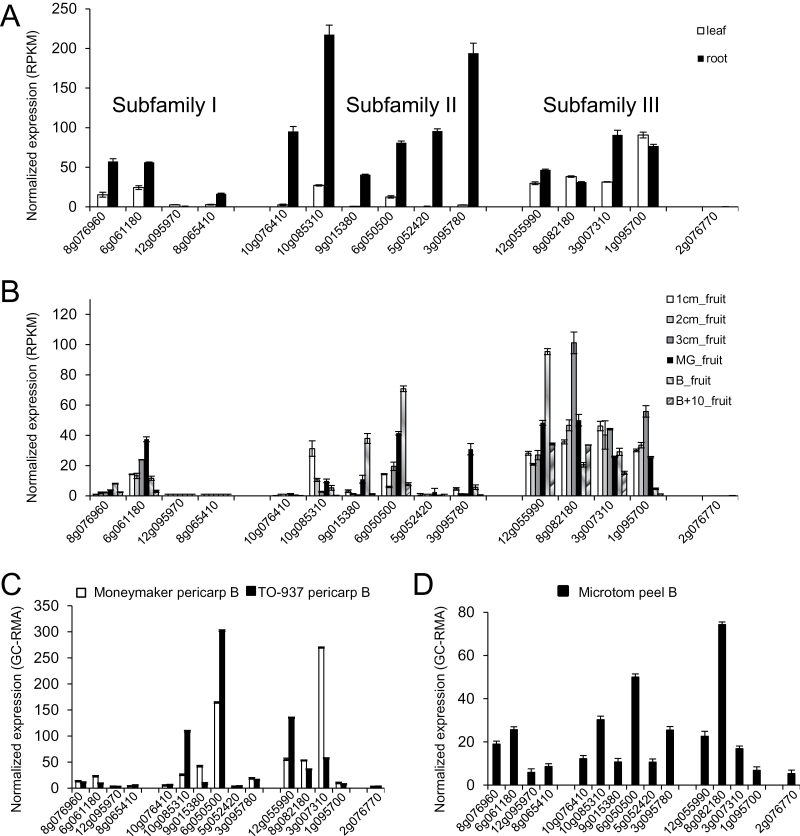
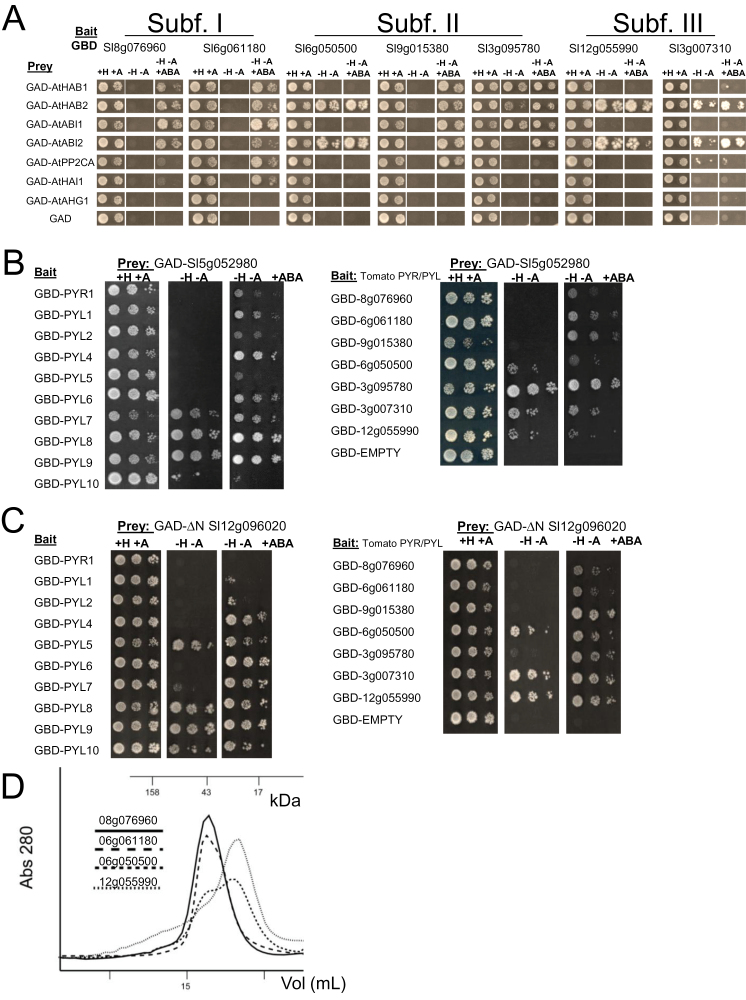
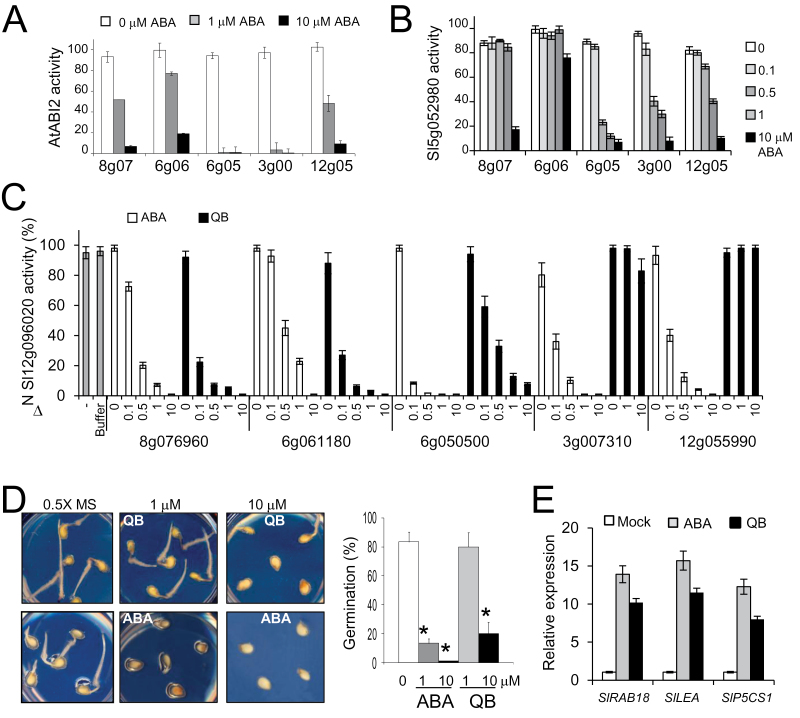
Abscisic acid (ABA) plays a crucial role in the plant's response to both biotic and abiotic stress. Sustainable production of food faces several key challenges, particularly the generation of new varieties with improved water use efficiency and drought tolerance. Different studies have shown the potential applications of Arabidopsis PYR/PYL/RCAR ABA receptors to enhance plant drought resistance. Consequently the functional characterization of orthologous genes in crops holds promise for agriculture. The full set of tomato (Solanum lycopersicum) PYR/PYL/RCAR ABA receptors have been identified here. From the 15 putative tomato ABA receptors, 14 of them could be grouped in three subfamilies that correlated well with corresponding Arabidopsis subfamilies. High levels of expression of PYR/PYL/RCAR genes was found in tomato root, and some genes showed predominant expression in leaf and fruit tissues. Functional characterization of tomato receptors was performed through interaction assays with Arabidopsis and tomato clade A protein phosphatase type 2Cs (PP2Cs) as well as phosphatase inhibition studies. Tomato receptors were able to inhibit the activity of clade A PP2Cs differentially in an ABA-dependent manner, and at least three receptors were sensitive to the ABA agonist quinabactin, which inhibited tomato seed germination. Indeed, the chemical activation of ABA signalling induced by quinabactin was able to activate stress-responsive genes. Both dimeric and monomeric tomato receptors were functional in Arabidopsis plant cells, but only overexpression of monomeric-type receptors conferred enhanced drought resistance. In summary, gene expression analyses, and chemical and transgenic approaches revealed distinct properties of tomato PYR/PYL/RCAR ABA receptors that might have biotechnological implications.
Keywords: Abscisic acid (ABA); drought resistance; tomato ABA receptor; tomato clade A PP2C..
© The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
Similar articles
-
Unique drought resistance functions of the highly ABA-induced clade A protein phosphatase 2Cs.Plant Physiol. 2012 Sep;160(1):379-95. doi: 10.1104/pp.112.202408. Epub 2012 Jul 24. Plant Physiol. 2012. PMID: 22829320 Free PMC article.
-
Structural insights into PYR/PYL/RCAR ABA receptors and PP2Cs.Plant Sci. 2012 Jan;182:3-11. doi: 10.1016/j.plantsci.2010.11.014. Epub 2010 Dec 7. Plant Sci. 2012. PMID: 22118610 Review.
-
Selective inhibition of clade A phosphatases type 2C by PYR/PYL/RCAR abscisic acid receptors.Plant Physiol. 2012 Feb;158(2):970-80. doi: 10.1104/pp.111.188623. Epub 2011 Dec 23. Plant Physiol. 2012. PMID: 22198272 Free PMC article.
-
PYR/PYL/RCAR family members are major in-vivo ABI1 protein phosphatase 2C-interacting proteins in Arabidopsis.Plant J. 2010 Jan;61(2):290-9. doi: 10.1111/j.1365-313X.2009.04054.x. Epub 2009 Oct 26. Plant J. 2010. PMID: 19874541 Free PMC article.
-
Early abscisic acid signal transduction mechanisms: newly discovered components and newly emerging questions.Genes Dev. 2010 Aug 15;24(16):1695-708. doi: 10.1101/gad.1953910. Genes Dev. 2010. PMID: 20713515 Free PMC article. Review.
Cited by
-
TaPYL4, an ABA receptor gene of wheat, positively regulates plant drought adaptation through modulating the osmotic stress-associated processes.BMC Plant Biol. 2022 Sep 1;22(1):423. doi: 10.1186/s12870-022-03799-z. BMC Plant Biol. 2022. PMID: 36050643 Free PMC article.
-
ABA signaling branches out: emerging ABA-related signaling functions in Solanum tuberosum.J Exp Bot. 2023 Nov 21;74(21):6405-6408. doi: 10.1093/jxb/erad395. J Exp Bot. 2023. PMID: 37988178 Free PMC article.
-
Genome-wide analysis of the PYL-PP2C-SnRK2s family in the ABA signaling pathway of pitaya reveals its expression profiles under canker disease stress.BMC Genomics. 2024 Aug 1;25(1):749. doi: 10.1186/s12864-024-10665-9. BMC Genomics. 2024. PMID: 39090531 Free PMC article.
-
Co-silencing of ABA receptors (SlRCAR) reveals interactions between ABA and ethylene signaling during tomato fruit ripening.Hortic Res. 2022 Jun 5;9:uhac057. doi: 10.1093/hr/uhac057. eCollection 2022. Hortic Res. 2022. PMID: 35685223 Free PMC article.
-
Abscisic Acid: Role in Fruit Development and Ripening.Front Plant Sci. 2022 May 10;13:817500. doi: 10.3389/fpls.2022.817500. eCollection 2022. Front Plant Sci. 2022. PMID: 35620694 Free PMC article. Review.
References
-
- Baykov AA, Evtushenko OA, Avaeva SM. 1988. A malachite green procedure for orthophosphate determination and its use in alkaline phosphatase-based enzyme immunoassay. Analytical Biochemistry 171, 266–270 - PubMed
-
- Ben-Ari G. 2012. The ABA signal transduction mechanism in commercial crops: learning from Arabidopsis . Plant Cell Reports 31, 1357–1369 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
