

Calmodulin regulation (calmodulation) of voltage-gated calcium channels
- PMID: 24863929
- PMCID: PMC4035741
- DOI: 10.1085/jgp.201311153
Calmodulin regulation (calmodulation) of voltage-gated calcium channels
Abstract
Calmodulin regulation (calmodulation) of the family of voltage-gated CaV1-2 channels comprises a prominent prototype for ion channel regulation, remarkable for its powerful Ca(2+) sensing capabilities, deep in elegant mechanistic lessons, and rich in biological and therapeutic implications. This field thereby resides squarely at the epicenter of Ca(2+) signaling biology, ion channel biophysics, and therapeutic advance. This review summarizes the historical development of ideas in this field, the scope and richly patterned organization of Ca(2+) feedback behaviors encompassed by this system, and the long-standing challenges and recent developments in discerning a molecular basis for calmodulation. We conclude by highlighting the considerable synergy between mechanism, biological insight, and promising therapeutics.
© 2014 Ben-Johny and Yue.
Figures
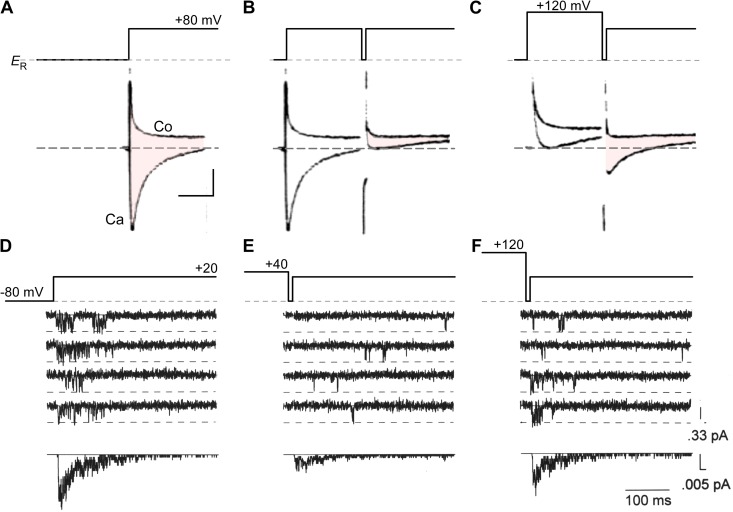
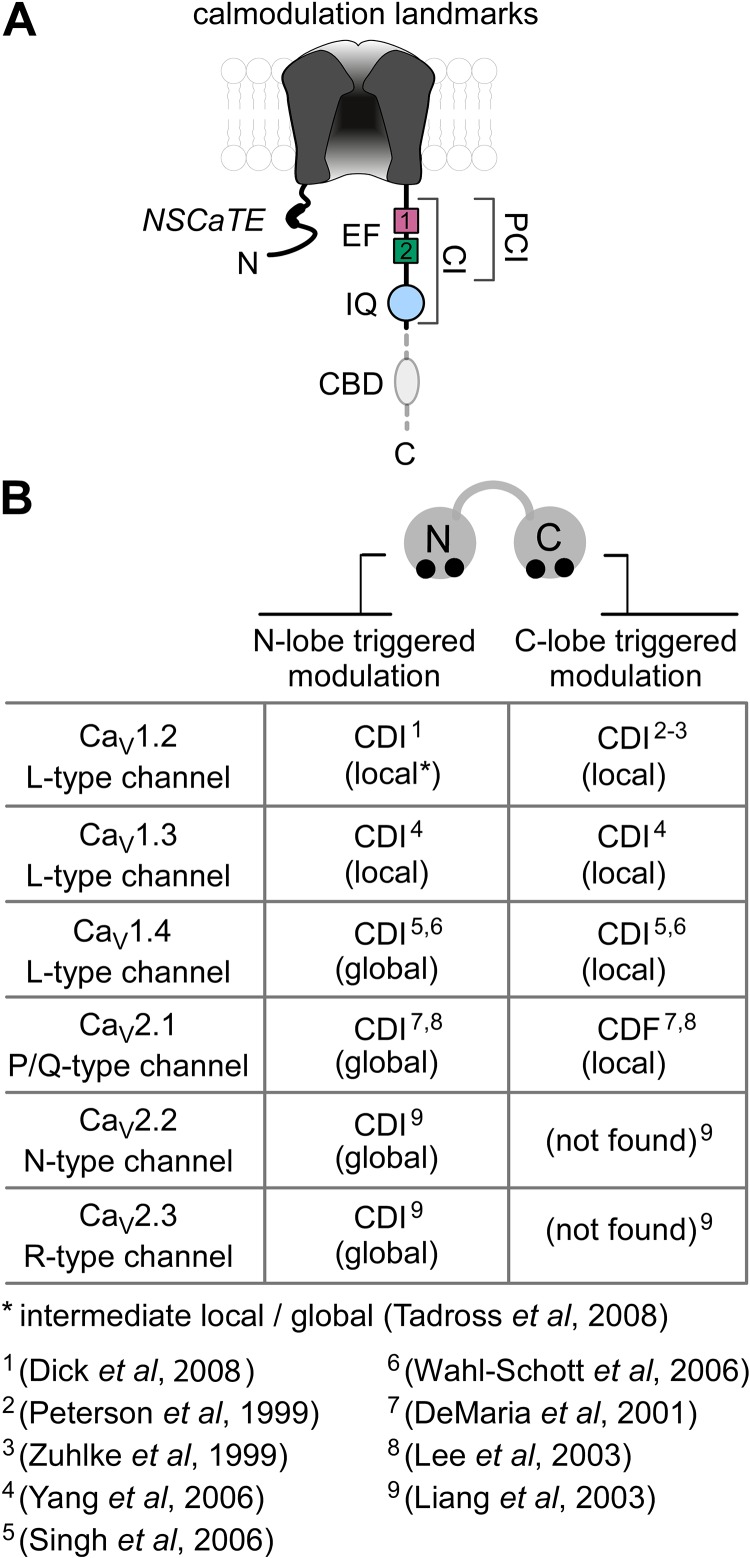
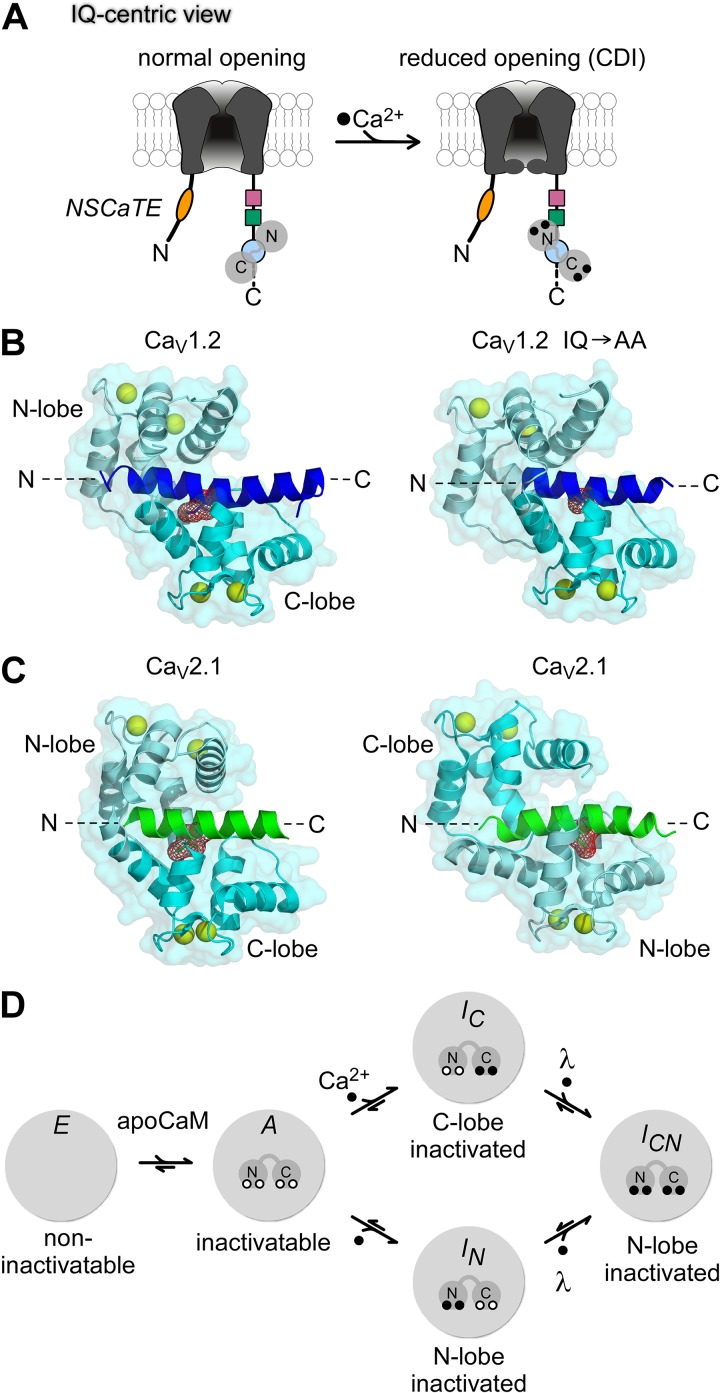

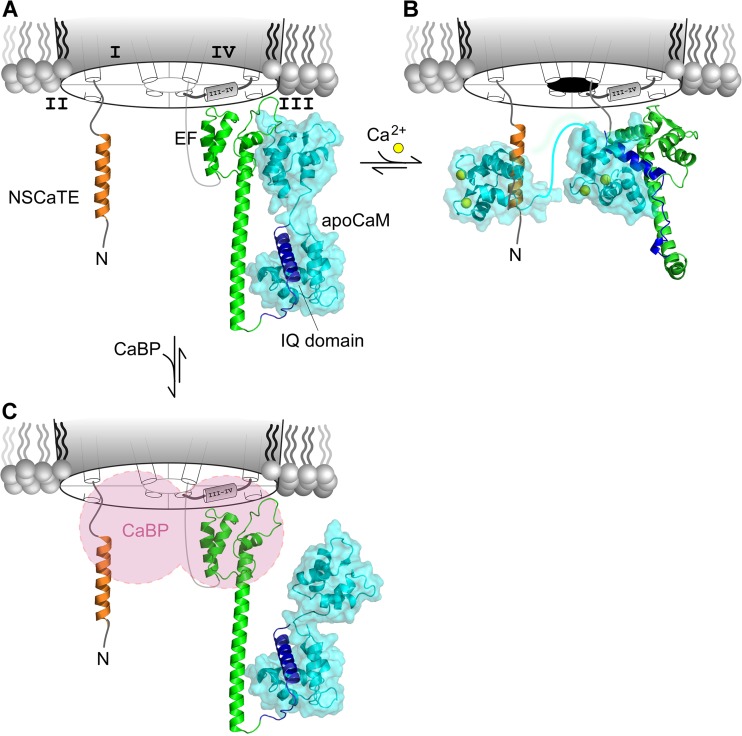
References
-
- Adams P.J., Rungta R.L., Garcia E., van den Maagdenberg A.M., MacVicar B.A., Snutch T.P. 2010. Contribution of calcium-dependent facilitation to synaptic plasticity revealed by migraine mutations in the P/Q-type calcium channel. Proc. Natl. Acad. Sci. USA. 107:18694–18699 10.1073/pnas.1009500107 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
