

Biochemical defects in minor spliceosome function in the developmental disorder MOPD I
- PMID: 24865609
- PMCID: PMC4114687
- DOI: 10.1261/rna.045187.114
Biochemical defects in minor spliceosome function in the developmental disorder MOPD I
Abstract
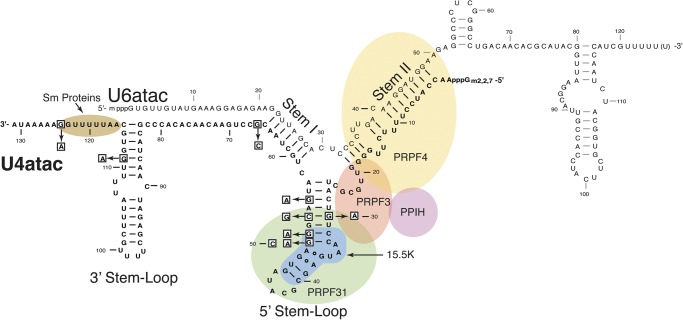
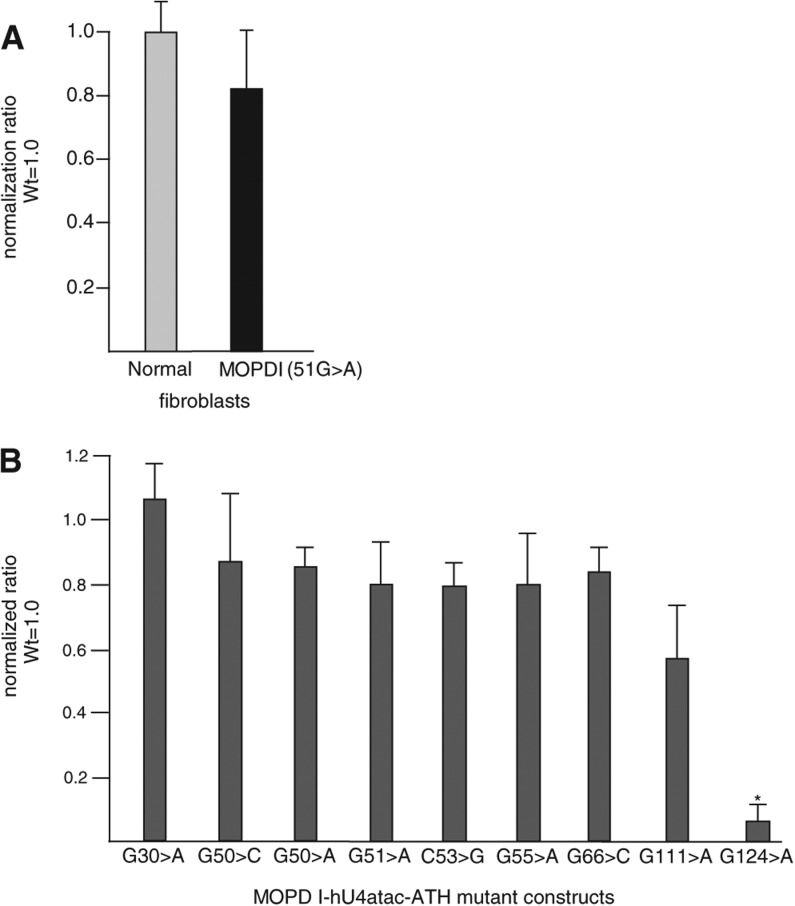
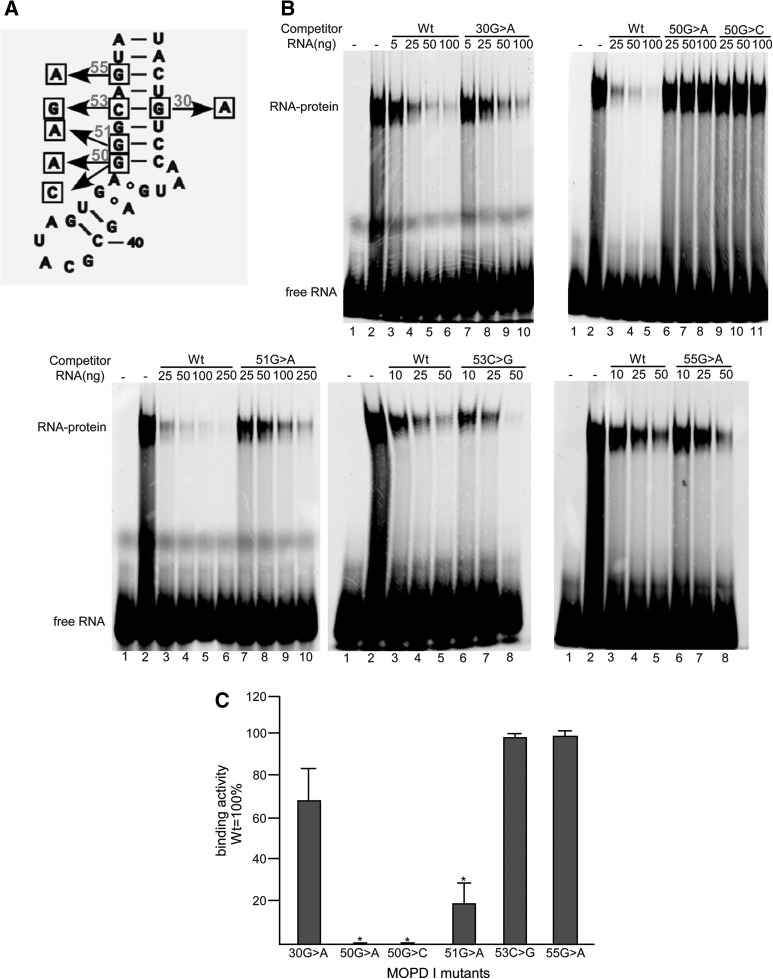
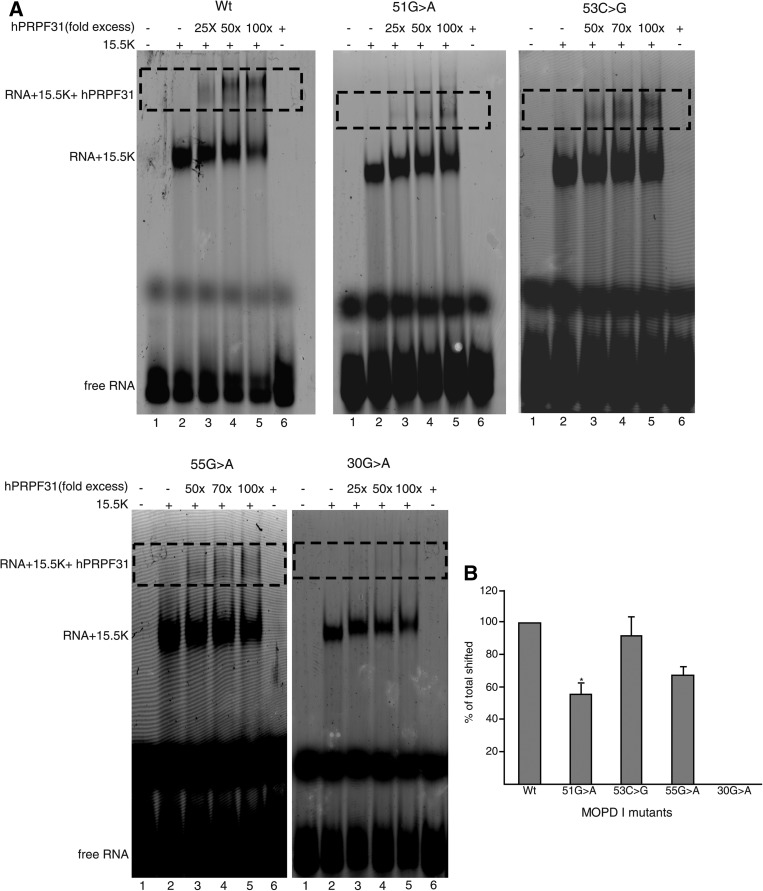
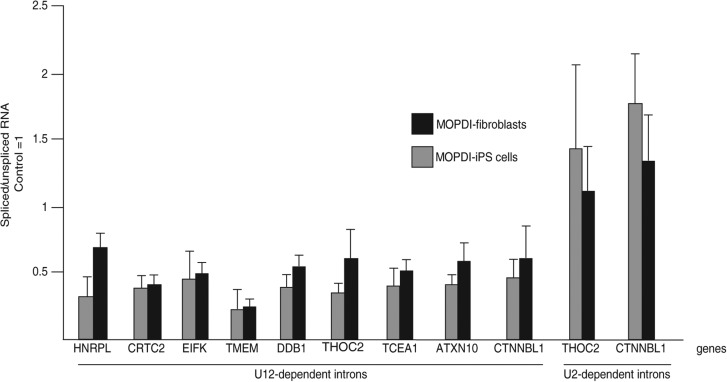
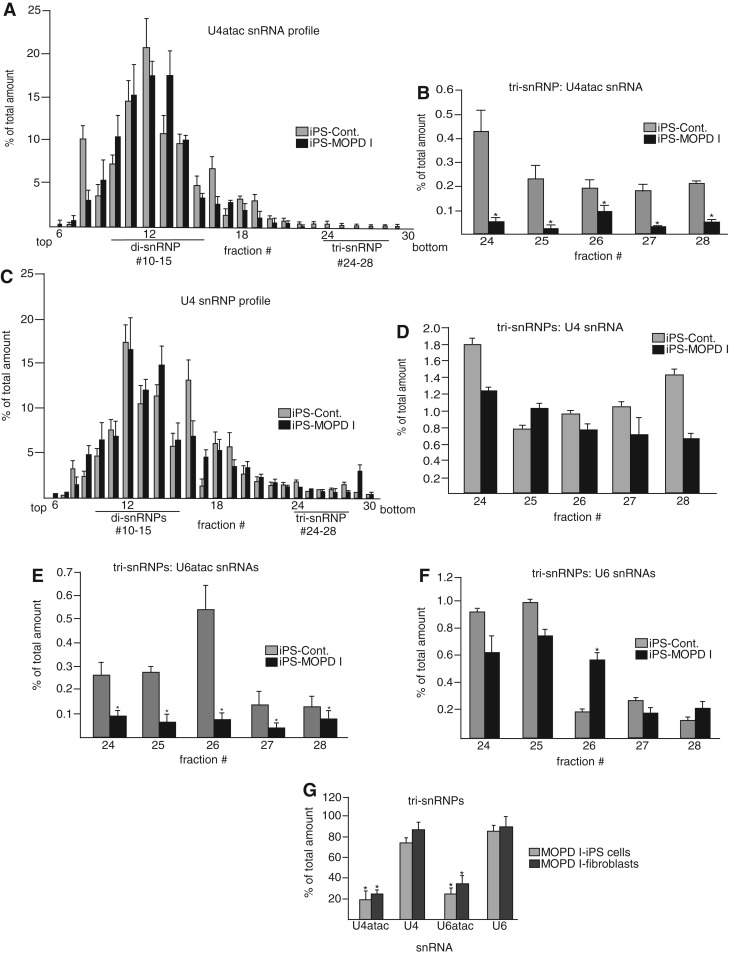
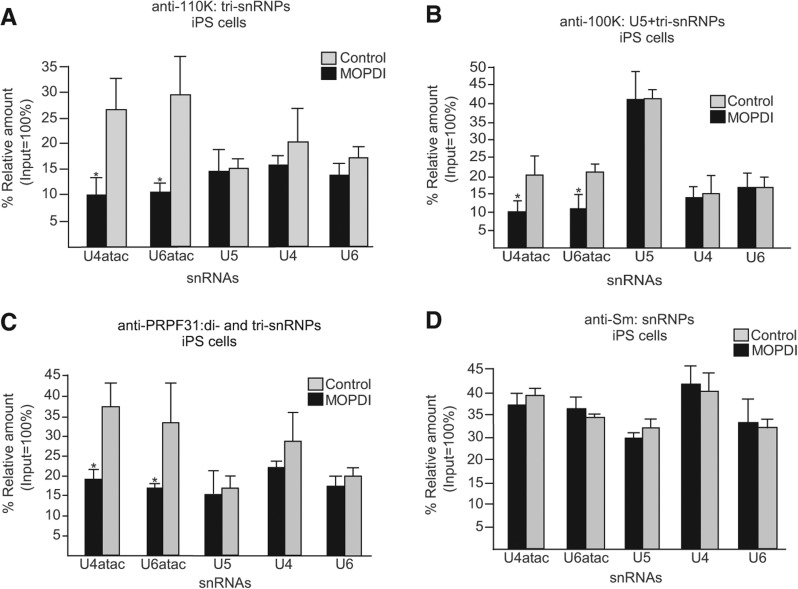
Biallelic mutations of the human RNU4ATAC gene, which codes for the minor spliceosomal U4atac snRNA, cause the developmental disorder, MOPD I/TALS. To date, nine separate mutations in RNU4ATAC have been identified in MOPD I patients. Evidence suggests that all of these mutations lead to abrogation of U4atac snRNA function and impaired minor intron splicing. However, the molecular basis of these effects is unknown. Here, we use a variety of in vitro and in vivo assays to address this question. We find that only one mutation, 124G>A, leads to significantly reduced expression of U4atac snRNA, whereas four mutations, 30G>A, 50G>A, 50G>C and 51G>A, show impaired binding of essential protein components of the U4atac/U6atac di-snRNP in vitro and in vivo. Analysis of MOPD I patient fibroblasts and iPS cells homozygous for the most common mutation, 51G>A, shows reduced levels of the U4atac/U6atac.U5 tri-snRNP complex as determined by glycerol gradient sedimentation and immunoprecipitation. In this report, we establish a mechanistic basis for MOPD I disease and show that the inefficient splicing of genes containing U12-dependent introns in patient cells is due to defects in minor tri-snRNP formation, and the MOPD I-associated RNU4ATAC mutations can affect multiple facets of minor snRNA function.
Keywords: U4atac snRNA; disease; snRNP function; splicing.
© 2014 Jafarifar et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
-
- Abdel-Salam GMH, Miyake N, Eid MM, Abdel-Hamid MS, Hassan NA, Eid OM, Effat LK, El-Badry TH, El-Kamah GY, El-Darouti M, et al. 2011. A homozygous mutation in RNU4ATAC as a cause of microcephalic osteodysplastic primordial dwarfism type I (MOPD I) with associated pigmentary disorder. Am J Med Genet A 155A: 2885–2896 - PubMed
-
- Abdel-Salam GM, Abdel-Hamid MS, Issa M, Magdy A, El-Kotoury A, Amr K 2012. Expanding the phenotypic and mutational spectrum in microcephalic osteodysplastic primordial dwarfism type I. Am J Med Genet A 158A: 1455–1461 - PubMed
-
- Abdel-Salam GM, Abdel-Hamid MS, Hassan NA, Issa MY, Effat L, Ismail S, Aglan MS, Zaki MS 2013. Further delineation of the clinical spectrum in RNU4ATAC related microcephalic osteodysplastic primordial dwarfism type I. Am J Med Genet A 161A: 1875–1881 - PubMed
-
- Boulisfane N, Choleza M, Rage F, Neel H, Soret J, Bordonné R 2011. Impaired minor tri-snRNP assembly generates differential splicing defects of U12-type introns in lymphoblasts derived from a type I SMA patient. Hum Mol Genet 20: 641–648 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical