

PZR coordinates Shp2 Noonan and LEOPARD syndrome signaling in zebrafish and mice
- PMID: 24865967
- PMCID: PMC4135572
- DOI: 10.1128/MCB.00135-14
PZR coordinates Shp2 Noonan and LEOPARD syndrome signaling in zebrafish and mice
Abstract
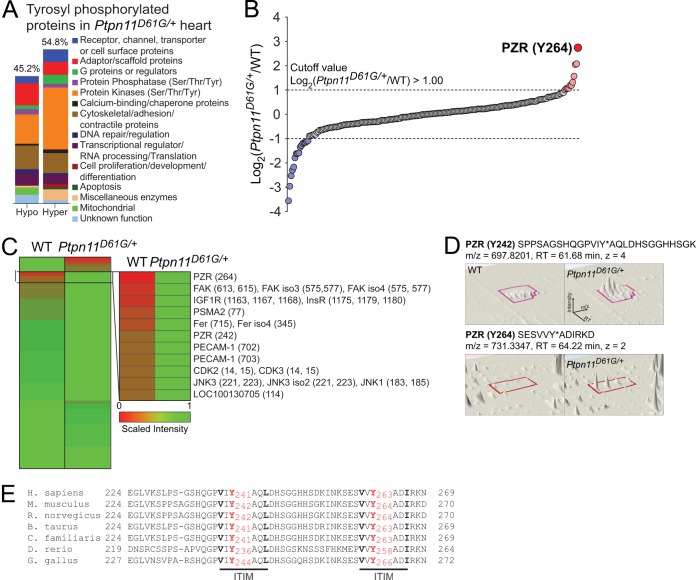
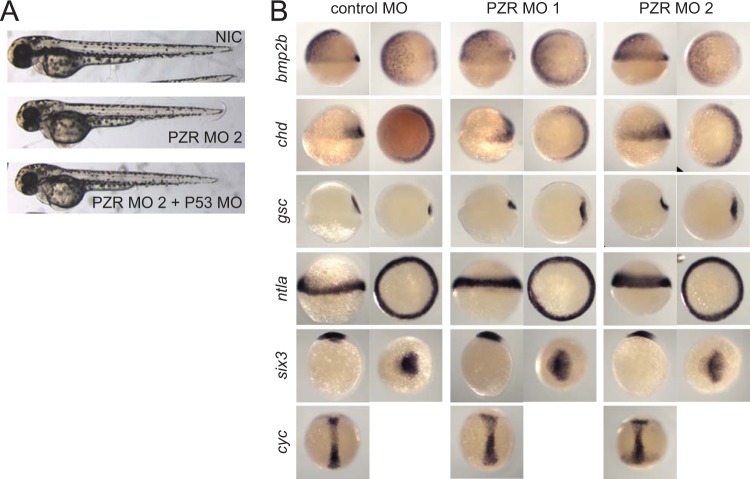
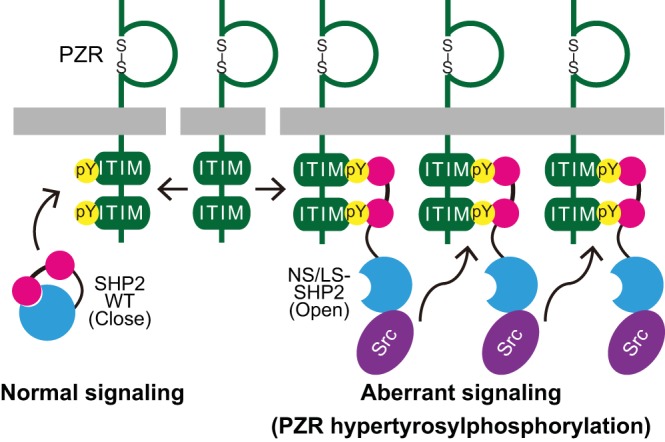
Noonan syndrome (NS) is an autosomal dominant disorder caused by activating mutations in the PTPN11 gene encoding Shp2, which manifests in congenital heart disease, short stature, and facial dysmorphia. The complexity of Shp2 signaling is exemplified by the observation that LEOPARD syndrome (LS) patients possess inactivating PTPN11 mutations yet exhibit similar symptoms to NS. Here, we identify "protein zero-related" (PZR), a transmembrane glycoprotein that interfaces with the extracellular matrix to promote cell migration, as a major hyper-tyrosyl-phosphorylated protein in mouse and zebrafish models of NS and LS. PZR hyper-tyrosyl phosphorylation is facilitated in a phosphatase-independent manner by enhanced Src recruitment to NS and LS Shp2. In zebrafish, PZR overexpression recapitulated NS and LS phenotypes. PZR was required for zebrafish gastrulation in a manner dependent upon PZR tyrosyl phosphorylation. Hence, we identify PZR as an NS and LS target. Enhanced PZR-mediated membrane recruitment of Shp2 serves as a common mechanism to direct overlapping pathophysiological characteristics of these PTPN11 mutations.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures











References
-
- Neel BG, Guo H, Pao L. 2009. SH2 domain-containing protein tyrosine phosphatases, p 707–728 In Bradshaw RA, Dennis EA. (ed), Handbook in cell signaling, 2nd ed, vol 2 Elsevier, San Diego, CA
-
- Mohi MG, Williams IR, Dearolf CR, Chan G, Kutok JL, Cohen S, Morgan K, Boulton C, Shigematsu H, Keilhack H, Akashi K, Gilliland DG, Neel BG. 2005. Prognostic, therapeutic, and mechanistic implications of a mouse model of leukemia evoked by Shp2 (PTPN11) mutations. Cancer Cell 7:179–191. 10.1016/j.ccr.2005.01.010 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous