

Switching on sex: transcriptional regulation of the testis-determining gene Sry
- PMID: 24866114
- PMCID: PMC4034426
- DOI: 10.1242/dev.107052
Switching on sex: transcriptional regulation of the testis-determining gene Sry
Abstract
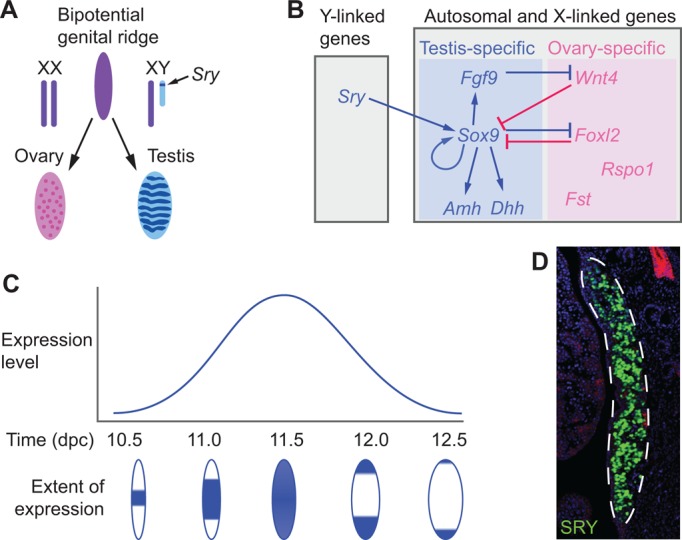
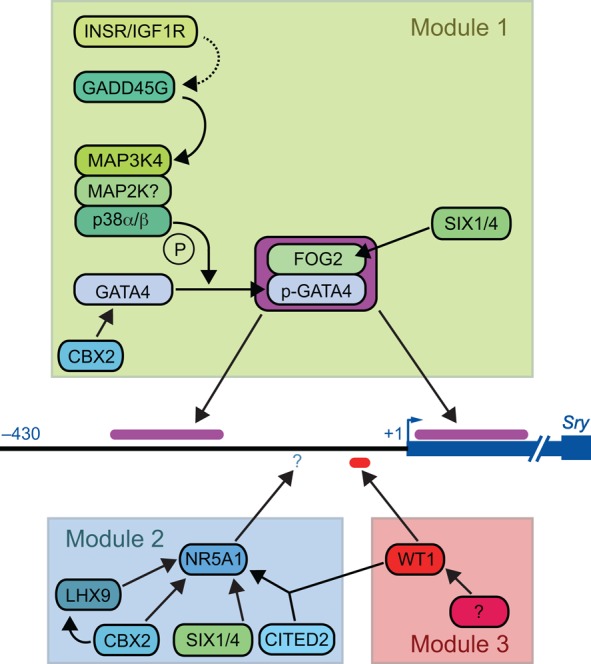
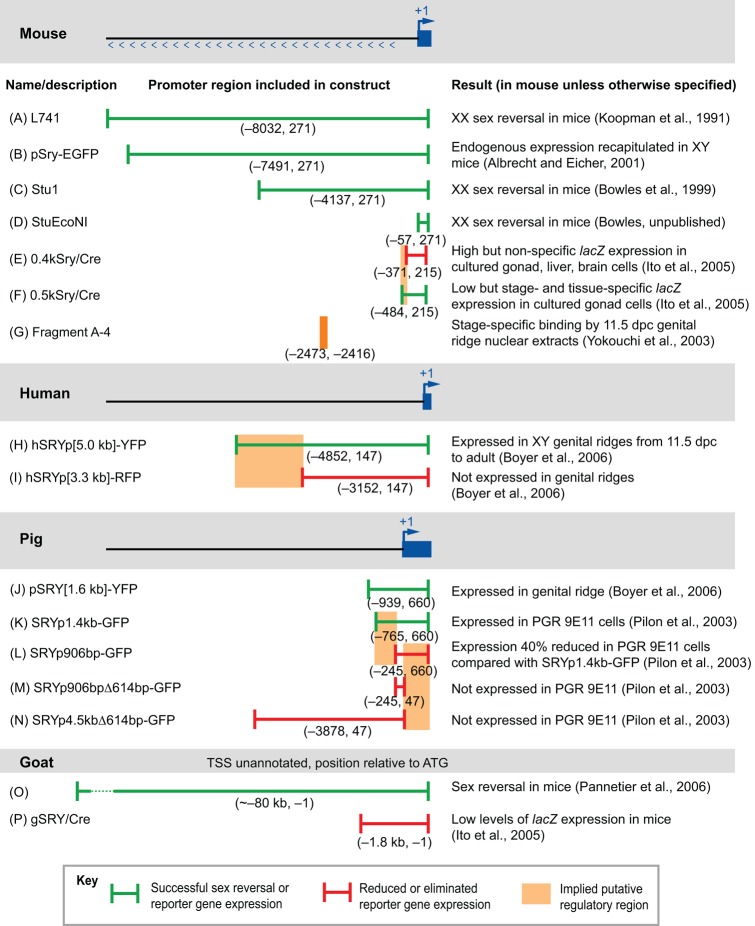
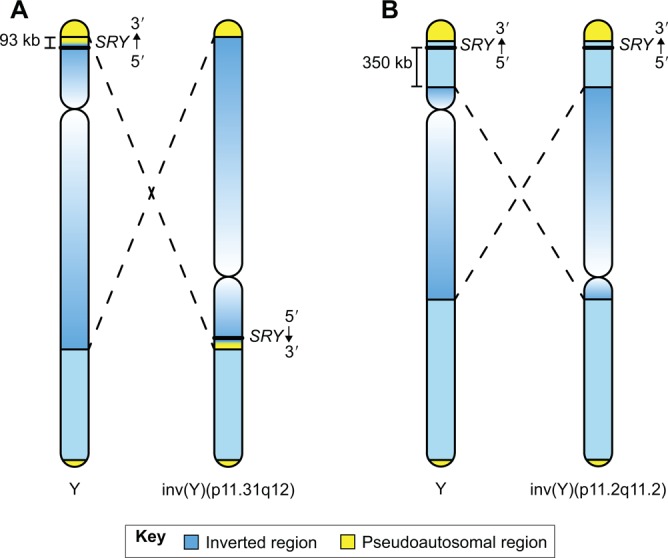
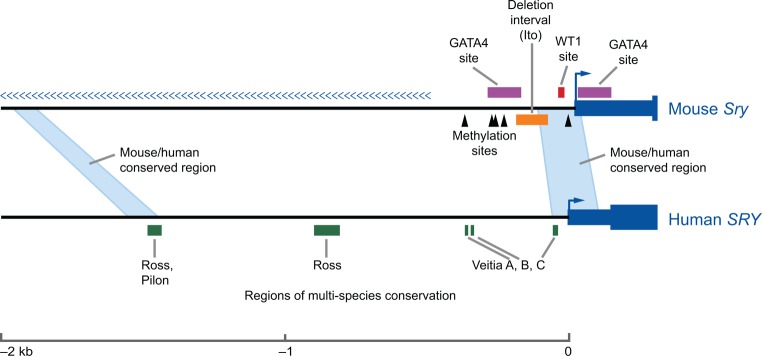
Mammalian sex determination hinges on the development of ovaries or testes, with testis fate being triggered by the expression of the transcription factor sex-determining region Y (Sry). Reduced or delayed Sry expression impairs testis development, highlighting the importance of its accurate spatiotemporal regulation and implying a potential role for SRY dysregulation in human intersex disorders. Several epigenetic modifiers, transcription factors and kinases are implicated in regulating Sry transcription, but it remains unclear whether or how this farrago of factors acts co-ordinately. Here we review our current understanding of Sry regulation and provide a model that assembles all known regulators into three modules, each converging on a single transcription factor that binds to the Sry promoter. We also discuss potential future avenues for discovering the cis-elements and trans-factors required for Sry regulation.
Keywords: Disorders of sex development; Sex determination; Sry; Testis development; Transcriptional regulation.
© 2014. Published by The Company of Biologists Ltd.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
