

SIRT2 interacts with β-catenin to inhibit Wnt signaling output in response to radiation-induced stress
- PMID: 24866770
- PMCID: PMC4163538
- DOI: 10.1158/1541-7786.MCR-14-0223-T
SIRT2 interacts with β-catenin to inhibit Wnt signaling output in response to radiation-induced stress
Abstract
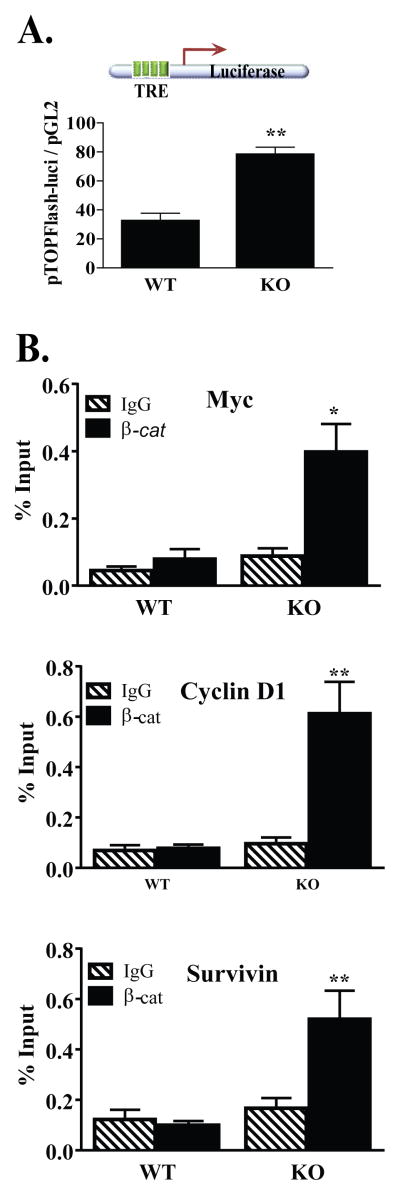
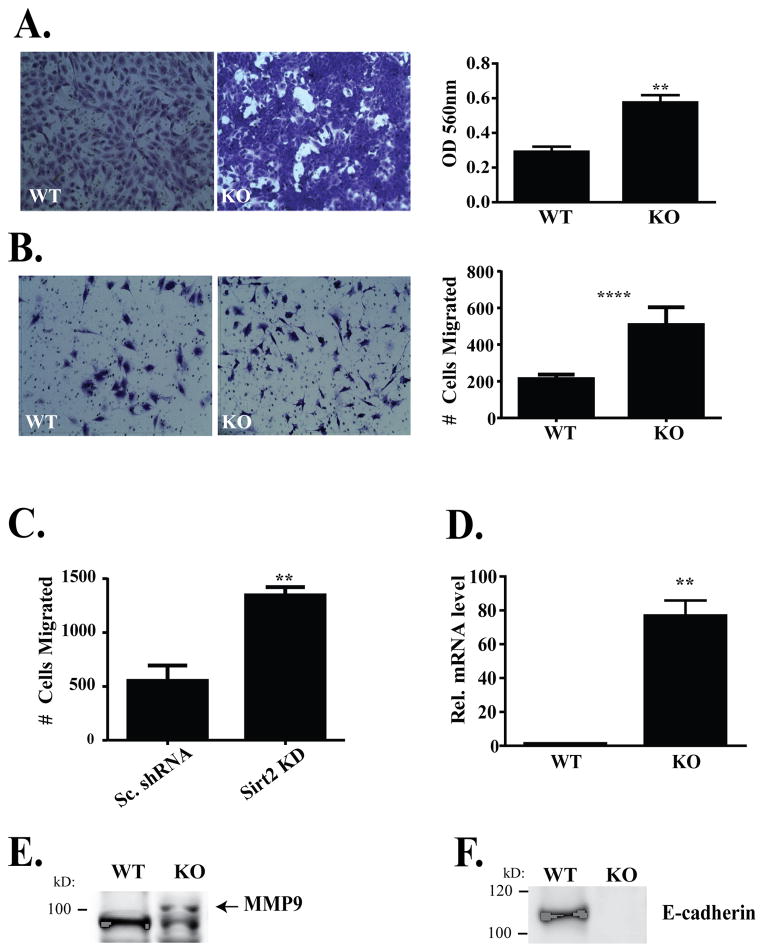
Wnt signaling is critical to maintaining cellular homeostasis via regulation of cell division, mitigation of cell stress, and degradation. Aberrations in Wnt signaling contribute to carcinogenesis and metastasis, whereas sirtuins have purported roles in carcinogenesis, aging, and neurodegeneration. Therefore, the hypothesis that sirtuin 2 (SIRT2) directly interacts with β-catenin and whether this interaction alters the expression of Wnt target genes to produce an altered cellular phenotype was tested. Coimmunoprecipitation studies, using mouse embryonic fibroblasts (MEF) from Sirt2 wild-type and genomic knockout mice, demonstrate that β-catenin directly binds SIRT2. Moreover, this interaction increases in response to oxidative stress induced by ionizing radiation. In addition, this association inhibits the expression of important Wnt target genes such as survivin (BIRC5), cyclin D1 (CCND1), and c-myc (MYC). In Sirt2 null MEFs, an upregulation of matrix metalloproteinase 9 (MMP9) and decreased E-cadherin (CDH1) expression is observed that produces increased cellular migration and invasion. Together, these data demonstrate that SIRT2, a tumor suppressor lost in multiple cancers, inhibits the Wnt signaling pathway in nonmalignant cells by binding to β-catenin and that SIRT2 plays a critical role in the response to oxidative stress from radiation.
Implications: Disruption of the SIRT2-β-catenin interaction represents an endogenous therapeutic target to prevent transformation and preserve the integrity of aging cells against exogenous stressors such as reactive oxygen species.
©2014 American Association for Cancer Research.
Conflict of interest statement
The authors disclose no potential conflicts of interest.
Figures
Similar articles
-
SIRT2 Contributes to the Regulation of Intestinal Cell Proliferation and Differentiation.Cell Mol Gastroenterol Hepatol. 2020;10(1):43-57. doi: 10.1016/j.jcmgh.2020.01.004. Epub 2020 Jan 16. Cell Mol Gastroenterol Hepatol. 2020. PMID: 31954883 Free PMC article.
-
Regulation of SIRT2 by Wnt/β-catenin signaling pathway in colorectal cancer cells.Biochim Biophys Acta Mol Cell Res. 2021 Apr;1868(4):118966. doi: 10.1016/j.bbamcr.2021.118966. Epub 2021 Jan 13. Biochim Biophys Acta Mol Cell Res. 2021. PMID: 33450304 Free PMC article.
-
Histone Demethylase JMJD2D Interacts With β-Catenin to Induce Transcription and Activate Colorectal Cancer Cell Proliferation and Tumor Growth in Mice.Gastroenterology. 2019 Mar;156(4):1112-1126. doi: 10.1053/j.gastro.2018.11.036. Epub 2018 Nov 23. Gastroenterology. 2019. PMID: 30472235
-
The role of Wnt/beta-catenin signaling in renal carcinogenesis: lessons from cadmium toxicity studies.Curr Mol Med. 2010 Jun;10(4):387-404. doi: 10.2174/156652410791316986. Curr Mol Med. 2010. PMID: 20455852 Review.
-
Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis.Front Immunol. 2018 Apr 13;9:745. doi: 10.3389/fimmu.2018.00745. eCollection 2018. Front Immunol. 2018. PMID: 29706964 Free PMC article. Review.
Cited by
-
SIRTain regulators of premature senescence and accelerated aging.Protein Cell. 2015 May;6(5):322-33. doi: 10.1007/s13238-015-0149-1. Epub 2015 Apr 25. Protein Cell. 2015. PMID: 25907989 Free PMC article. Review.
-
Human Sirtuin 2 Localization, Transient Interactions, and Impact on the Proteome Point to Its Role in Intracellular Trafficking.Mol Cell Proteomics. 2016 Oct;15(10):3107-3125. doi: 10.1074/mcp.M116.061333. Epub 2016 Aug 8. Mol Cell Proteomics. 2016. PMID: 27503897 Free PMC article.
-
Role of Sirtuins in Tumor Angiogenesis.Front Oncol. 2020 Jan 17;9:1516. doi: 10.3389/fonc.2019.01516. eCollection 2019. Front Oncol. 2020. PMID: 32010617 Free PMC article. Review.
-
SIRT2 Contributes to the Regulation of Intestinal Cell Proliferation and Differentiation.Cell Mol Gastroenterol Hepatol. 2020;10(1):43-57. doi: 10.1016/j.jcmgh.2020.01.004. Epub 2020 Jan 16. Cell Mol Gastroenterol Hepatol. 2020. PMID: 31954883 Free PMC article.
-
Exosomal miR-130a-3p regulates osteogenic differentiation of Human Adipose-Derived stem cells through mediating SIRT7/Wnt/β-catenin axis.Cell Prolif. 2020 Oct;53(10):e12890. doi: 10.1111/cpr.12890. Epub 2020 Aug 17. Cell Prolif. 2020. PMID: 32808361 Free PMC article.
References
-
- Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004;20:781–810. - PubMed
-
- Gordon MD, Nusse R. Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem. 2006;281(32):22429–33. - PubMed
-
- Reya T, Clevers H. Wnt signalling in stem cells and cancer. Nature. 2005;434(7035):843–50. - PubMed
-
- Daniels DL, Weis WI. Beta-catenin directly displaces Groucho/TLE repressors from Tcf/Lef in Wnt-mediated transcription activation. Nat Struct Mol Biol. 2005;12(4):364–71. - PubMed
-
- Brack AS, Conboy MJ, Roy S, Lee M, Kuo CJ, Keller C, et al. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science. 2007;317(5839):807–10. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
