

Overexpression of pigeonpea stress-induced cold and drought regulatory gene (CcCDR) confers drought, salt, and cold tolerance in Arabidopsis
- PMID: 24868035
- PMCID: PMC4144763
- DOI: 10.1093/jxb/eru224
Overexpression of pigeonpea stress-induced cold and drought regulatory gene (CcCDR) confers drought, salt, and cold tolerance in Arabidopsis
Abstract
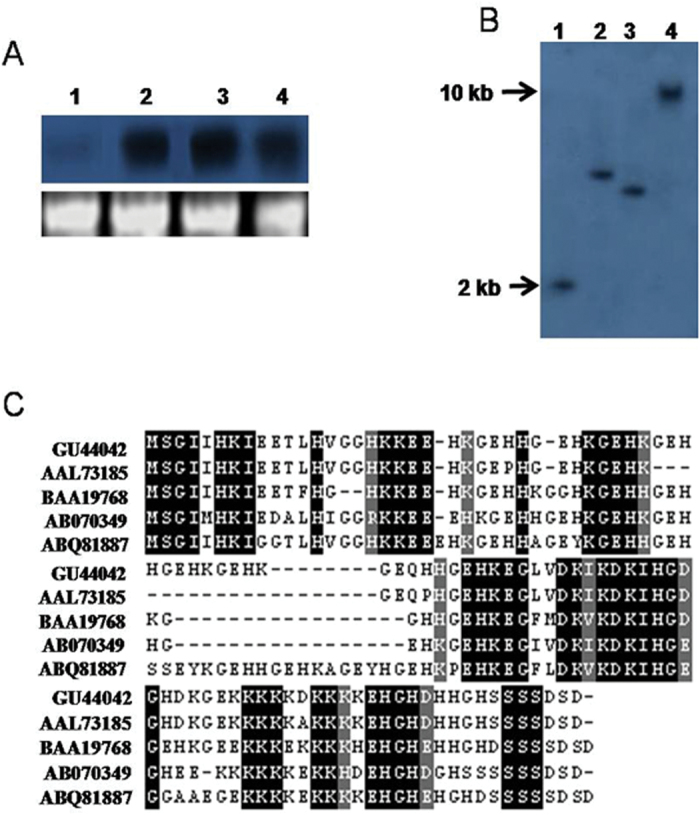
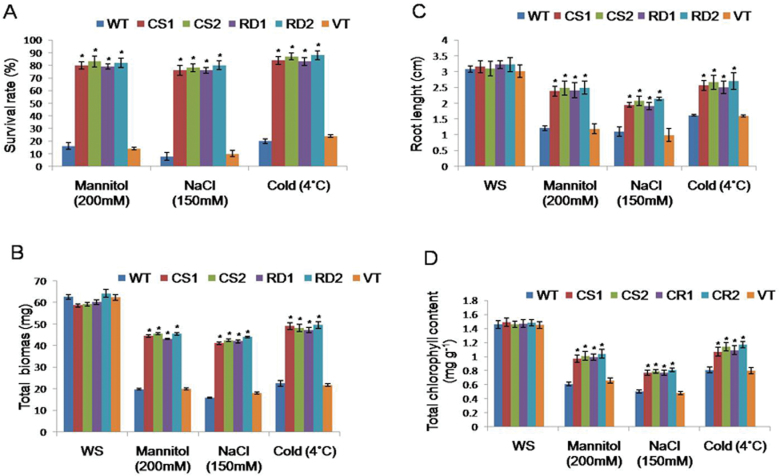
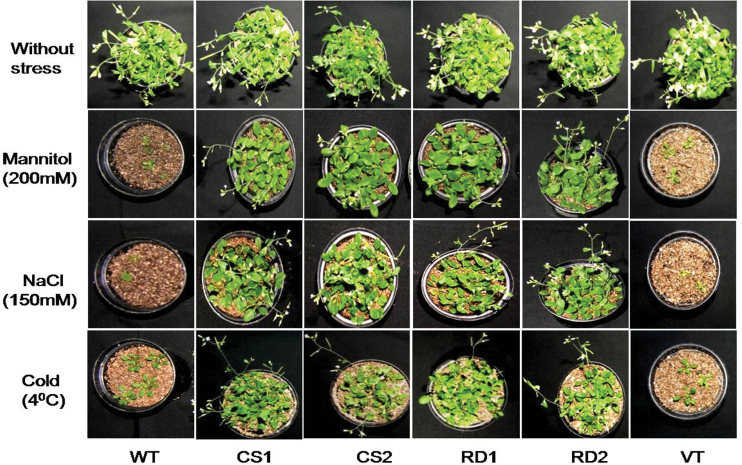
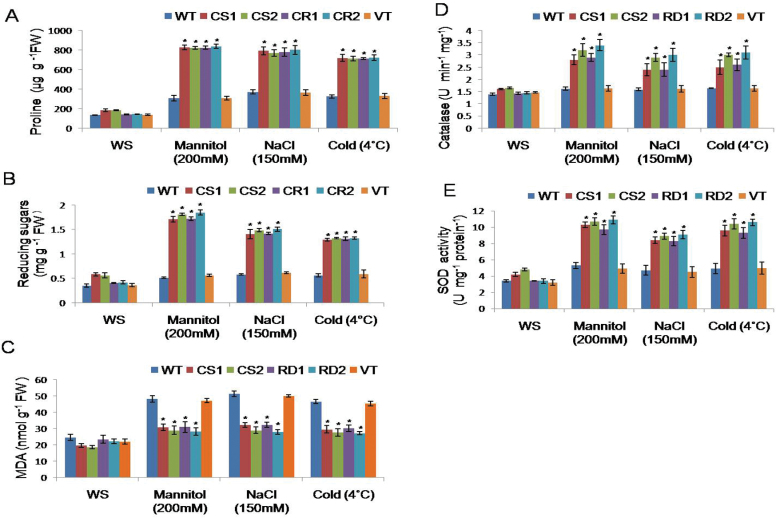
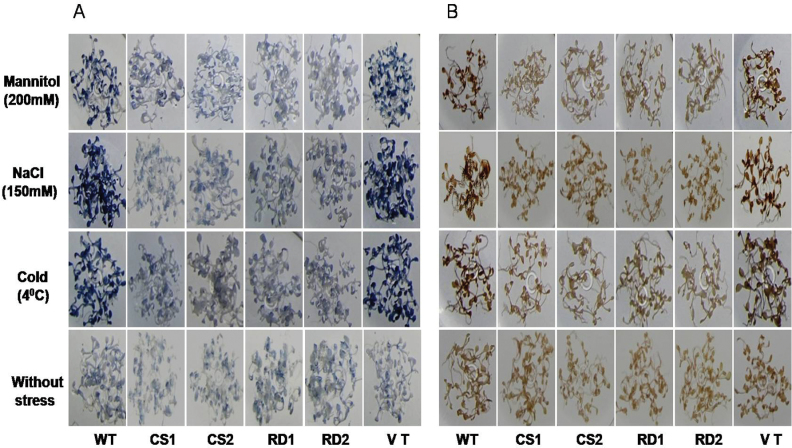
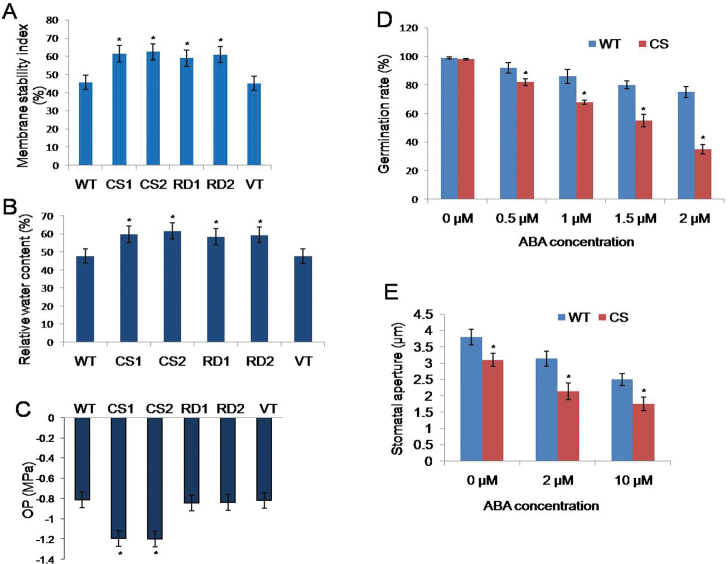
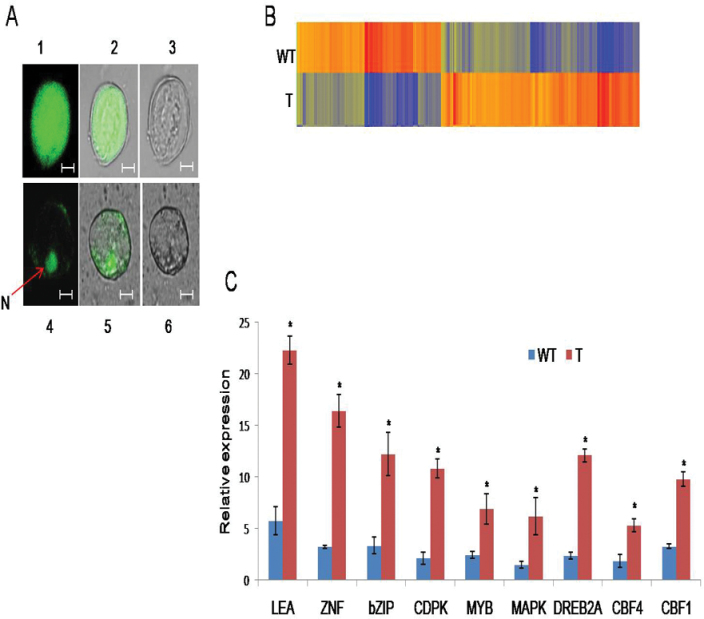
A potent cold and drought regulatory protein-encoding gene (CcCDR) was isolated from the subtractive cDNA library of pigeonpea plants subjected to drought stress. CcCDR was induced by different abiotic stress conditions in pigeonpea. Overexpression of CcCDR in Arabidopsis thaliana imparted enhanced tolerance against major abiotic stresses, namely drought, salinity, and low temperature, as evidenced by increased biomass, root length, and chlorophyll content. Transgenic plants also showed increased levels of antioxidant enzymes, proline, and reducing sugars under stress conditions. Furthermore, CcCDR-transgenic plants showed enhanced relative water content, osmotic potential, and cell membrane stability, as well as hypersensitivity to abscisic acid (ABA) as compared with control plants. Localization studies confirmed that CcCDR could enter the nucleus, as revealed by intense fluorescence, indicating its possible interaction with various nuclear proteins. Microarray analysis revealed that 1780 genes were up-regulated in CcCDR-transgenics compared with wild-type plants. Real-time PCR analysis on selected stress-responsive genes, involved in ABA-dependent and -independent signalling networks, revealed higher expression levels in transgenic plants, suggesting that CcCDR acts upstream of these genes. The overall results demonstrate the explicit role of CcCDR in conferring multiple abiotic stress tolerance at the whole-plant level. The multifunctional CcCDR seems promising as a prime candidate gene for enhancing abiotic stress tolerance in diverse plants.
Keywords: Abiotic stress tolerance; Cajanus cajan; cDNA library; cold and drought regulatory gene; nuclear localization..
© The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Apel K, Hirt H. 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology 55, 373–399 - PubMed
-
- Asano T, Hayashi N, Kobayashi M, et al. 2012. A rice calcium-dependent protein kinase OsCPK12 oppositely modulates salt-stress tolerance and blast disease resistance. The Plant Journal 69, 26–36 - PubMed
-
- Bates LS, Waldran R, Teare ID. 1973. Rapid determination of free proline for water studies. Plant and Soil 39, 205–208
-
- Brini F, Hanin M, Mezghani I, Berkowitz GA, Masmoudi K. 2007. Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphatase TVP1 improve salt- and drought stress tolerance in Arabidopsis thaliana plants. Journal of Experimental Botany 58, 301–308 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
