

Expression of actin-binding proteins and requirement for actin-depolymerizing factor in chick neural crest cells
- PMID: 24868596
- PMCID: PMC4048876
- DOI: 10.1002/dvdy.24105
Expression of actin-binding proteins and requirement for actin-depolymerizing factor in chick neural crest cells
Abstract
Background: Neural crest cells are multipotent cells that migrate extensively throughout vertebrate embryos to form diverse lineages. Cell migration requires polarized, organized actin networks that provide the driving force for motility. Actin-binding proteins that regulate neural crest cell migration are just beginning to be defined.
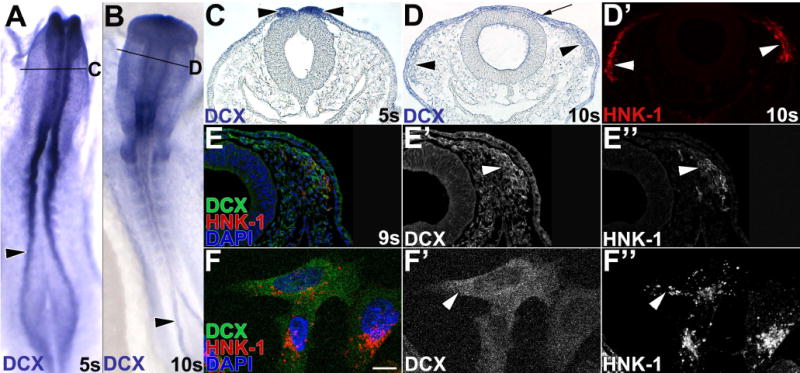
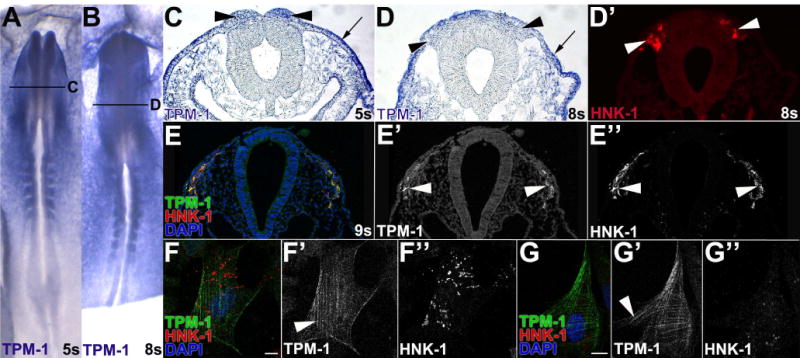
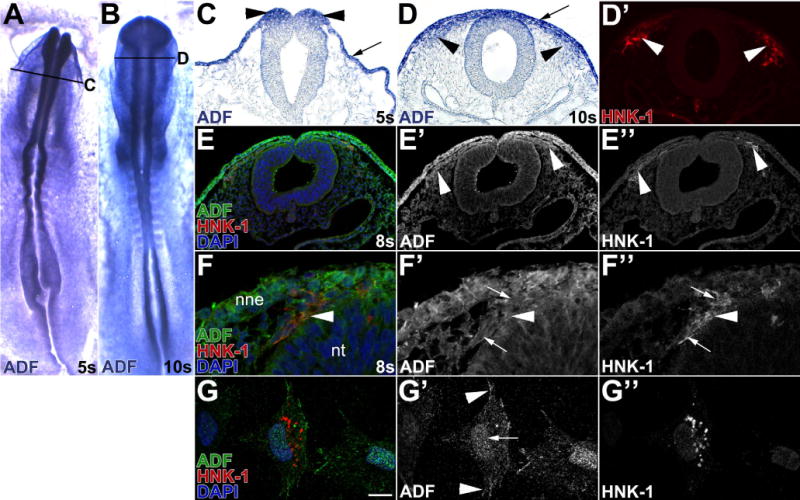
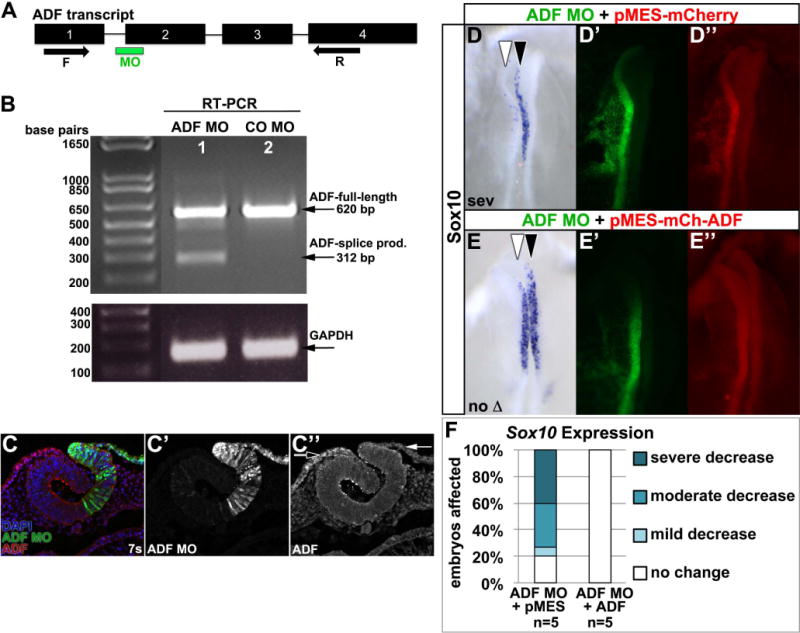
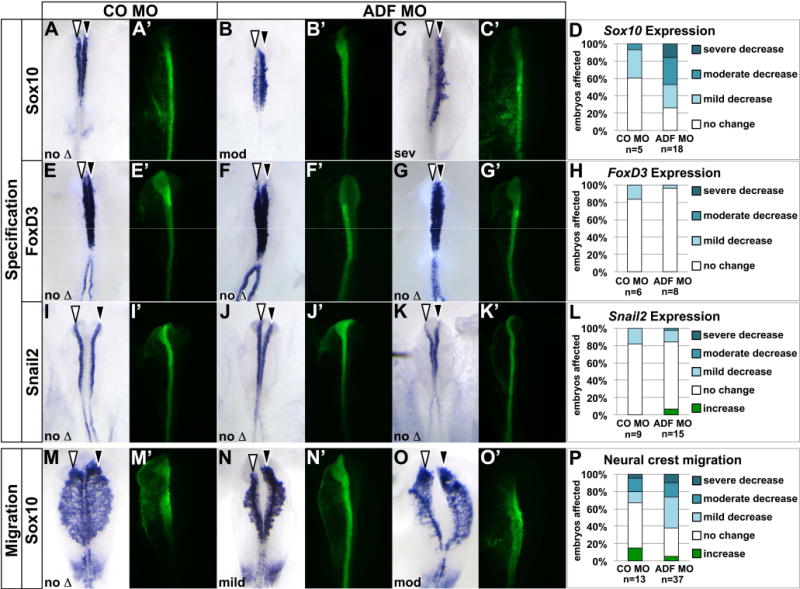
Results: We recently identified a number of actin-associated factors through proteomic profiling of methylated proteins in migratory neural crest cells. Here, we report the previously undocumented expression pattern of three of these proteins in chick early neural crest development: doublecortin (DCX), tropomyosin-1 (TPM-1), and actin depolymerizing factor (ADF). All three genes are expressed with varying degrees of specificity and intensity in premigratory and migratory neural crest cells, and their resulting proteins exhibit distinct subcellular localization in migratory neural crest cells. Morpholino knock down of ADF reveals it is required for Sox10 gene expression, but minimally important during neural crest migration.
Conclusions: Neural crest cells express DCX, TPM-1, and ADF. ADF is necessary during neural crest specification, but largely dispensable for migration.
Copyright © 2013 Wiley Periodicals, Inc.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
