

Orbitofrontal dopamine depletion upregulates caudate dopamine and alters behavior via changes in reinforcement sensitivity
- PMID: 24872570
- PMCID: PMC4035526
- DOI: 10.1523/JNEUROSCI.0718-14.2014
Orbitofrontal dopamine depletion upregulates caudate dopamine and alters behavior via changes in reinforcement sensitivity
Abstract
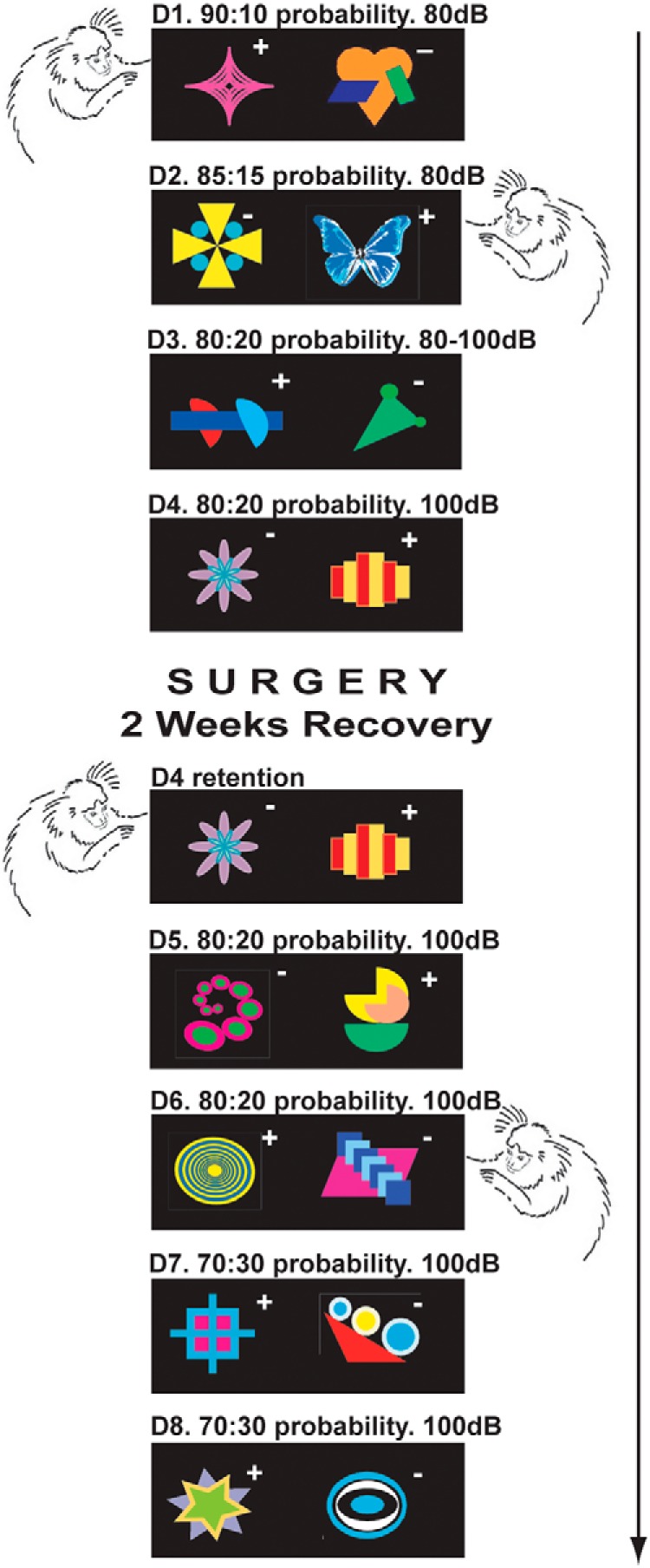
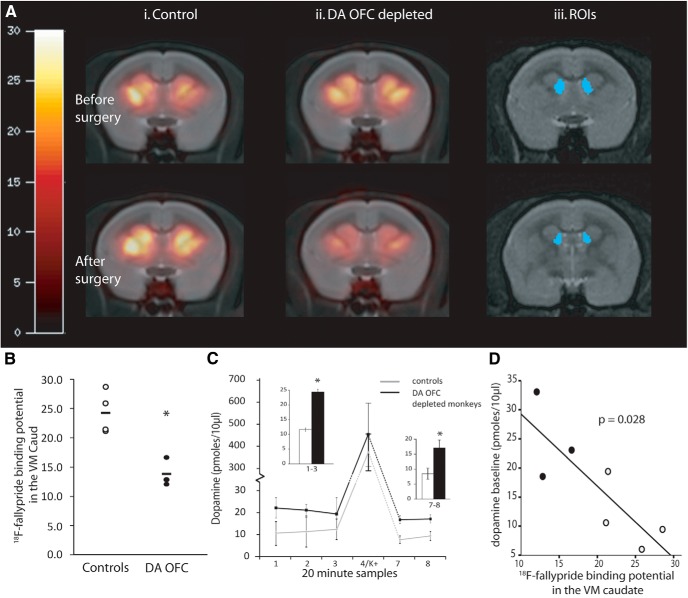
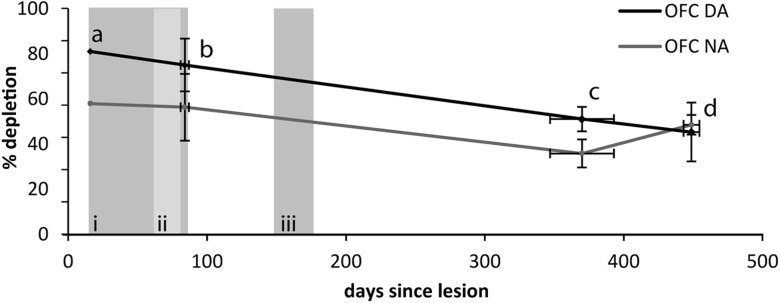
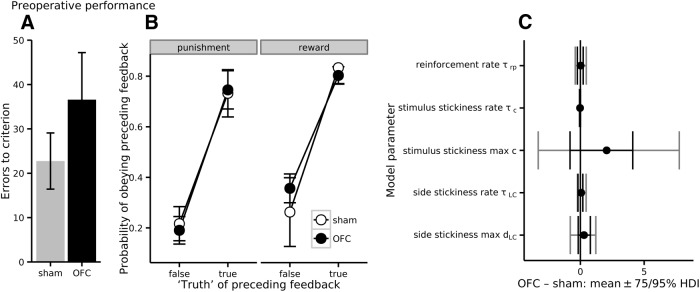
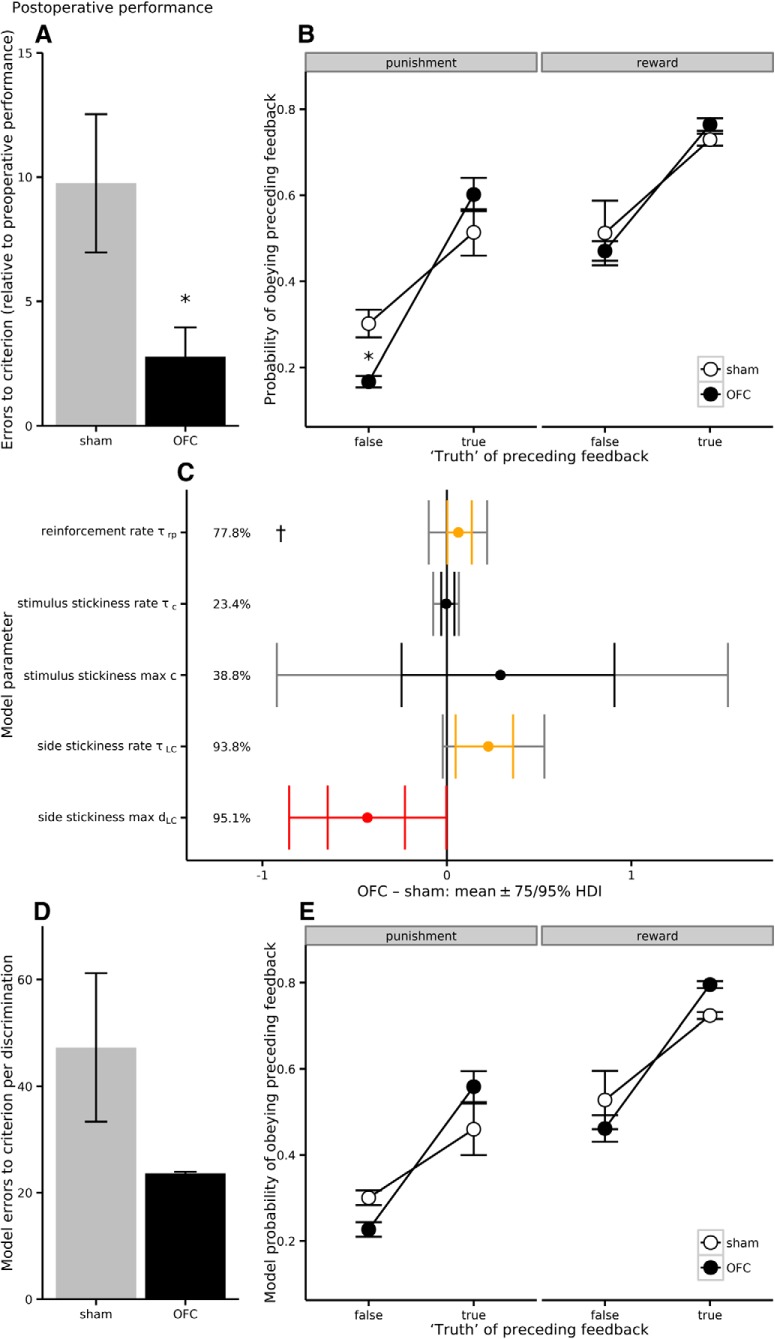
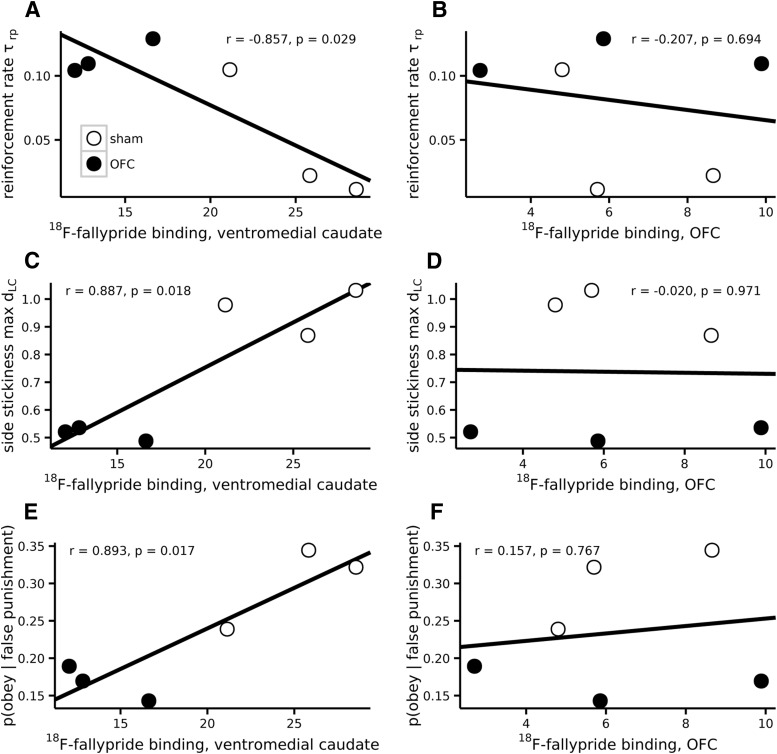
Schizophrenia is associated with upregulation of dopamine (DA) release in the caudate nucleus. The caudate has dense connections with the orbitofrontal cortex (OFC) via the frontostriatal loops, and both areas exhibit pathophysiological change in schizophrenia. Despite evidence that abnormalities in dopaminergic neurotransmission and prefrontal cortex function co-occur in schizophrenia, the influence of OFC DA on caudate DA and reinforcement processing is poorly understood. To test the hypothesis that OFC dopaminergic dysfunction disrupts caudate dopamine function, we selectively depleted dopamine from the OFC of marmoset monkeys and measured striatal extracellular dopamine levels (using microdialysis) and dopamine D2/D3 receptor binding (using positron emission tomography), while modeling reinforcement-related behavior in a discrimination learning paradigm. OFC dopamine depletion caused an increase in tonic dopamine levels in the caudate nucleus and a corresponding reduction in D2/D3 receptor binding. Computational modeling of behavior showed that the lesion increased response exploration, reducing the tendency to persist with a recently chosen response side. This effect is akin to increased response switching previously seen in schizophrenia and was correlated with striatal but not OFC D2/D3 receptor binding. These results demonstrate that OFC dopamine depletion is sufficient to induce striatal hyperdopaminergia and changes in reinforcement learning relevant to schizophrenia.
Keywords: PET; behavior; caudate nucleus; dopamine; orbitofrontal cortex; schizophrenia.
Copyright © 2014 Clarke, Cardinal et al.
Figures
References
-
- Bertolino A, Knable MB, Saunders RC, Callicott JH, Kolachana B, Mattay VS, Bachevalier J, Frank JA, Egan M, Weinberger DR. The relationship between dorsolateral prefrontal N-acetylaspartate measures and striatal dopamine activity in schizophrenia. Biol Psychiatry. 1999;45:660–667. doi: 10.1016/S0006-3223(98)00380-1. - DOI - PubMed
-
- Burnham KP, Anderson DR. Multimodel inference: understanding AIC and BIC in model selection. Sociol Methods Res. 2004;33:261–304. doi: 10.1177/0049124104268644. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources