

Regulation of nonmuscle myosin II by tropomyosin
- PMID: 24873380
- PMCID: PMC4075986
- DOI: 10.1021/bi500162z
Regulation of nonmuscle myosin II by tropomyosin
Abstract
The actin cytoskeleton carries out cellular functions, including division, migration, adhesion, and intracellular transport, that require a variety of actin binding proteins, including myosins. Our focus here is on class II nonmuscle myosin isoforms, NMIIA, NMIIB, and NMIIC, and their regulation by the actin binding protein, tropomyosin. NMII myosins are localized to different populations of stress fibers and the contractile ring, structures involved in force generation required for cell migration, adhesion, and cytokinesis. The stress fibers and contractile ring that contain NMII myosins also contain tropomyosin. Four mammalian genes encode more than 40 tropomyosins. Tropomyosins inhibit or activate actomyosin MgATPase and motility depending on the myosin and tropomyosin isoform. In vivo, tropomyosins play a role in cell migration, adhesion, cytokinesis, and NMII isoform localization in an isoform-specific manner. We postulate that the isoform-specific tropomyosin localization and effect on NMII isoform localization reflect modulation of NMII actomyosin kinetics and motile function. In this study, we compare the ability of different tropomyosin isoforms to support actin filament motility with NMIIA, NMIIB, and NMIIC as well as skeletal muscle myosin. Tropomyosins activated, inhibited, or had no effect on motility depending on the myosin, indicating that the myosin isoform is the primary determinant of the isoform-specific effect of tropomyosin on actomyosin regulation. Activation of motility of nonmuscle tropomyosin-actin filaments by NMII myosin correlates with an increased Vmax of the myosin MgATPase, implying a direct effect on the myosin MgATPase, in contrast to the skeletal tropomyosin-actin filament that has no effect on the Vmax or maximal filament velocity.
Figures
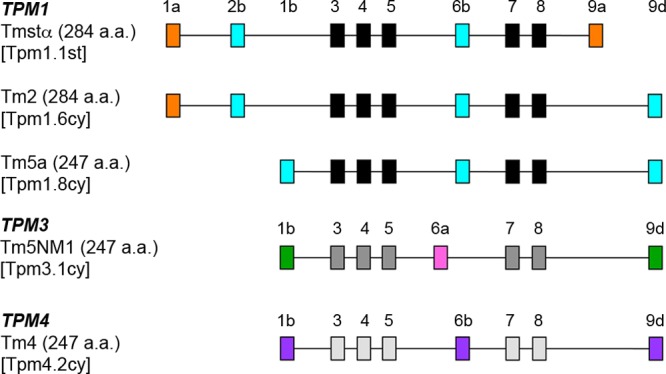
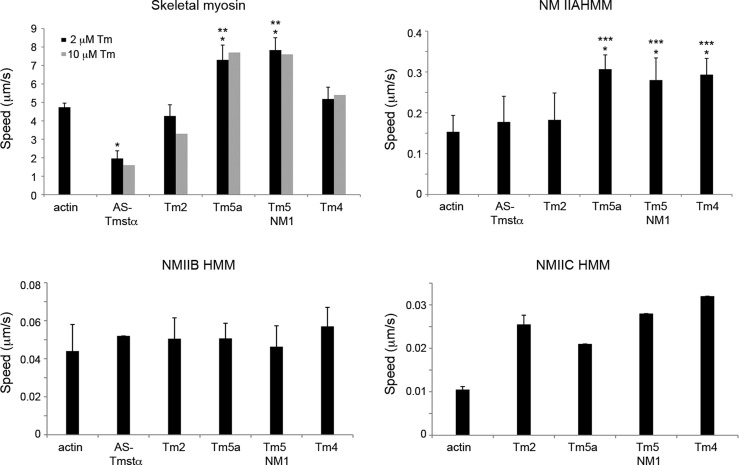
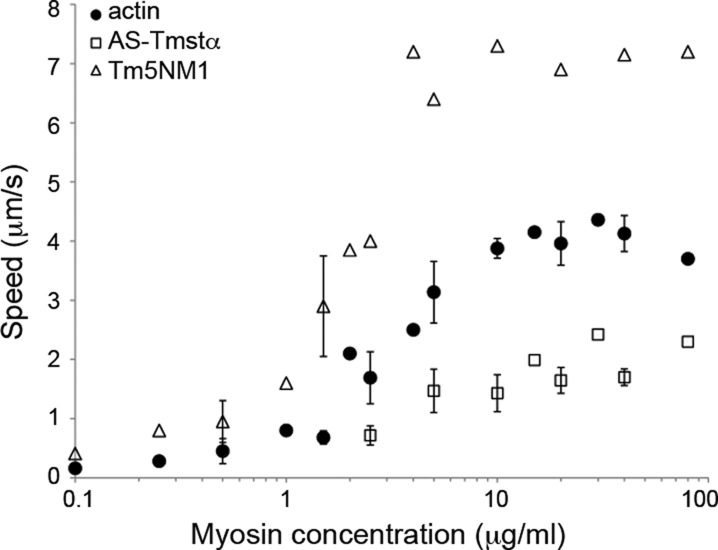
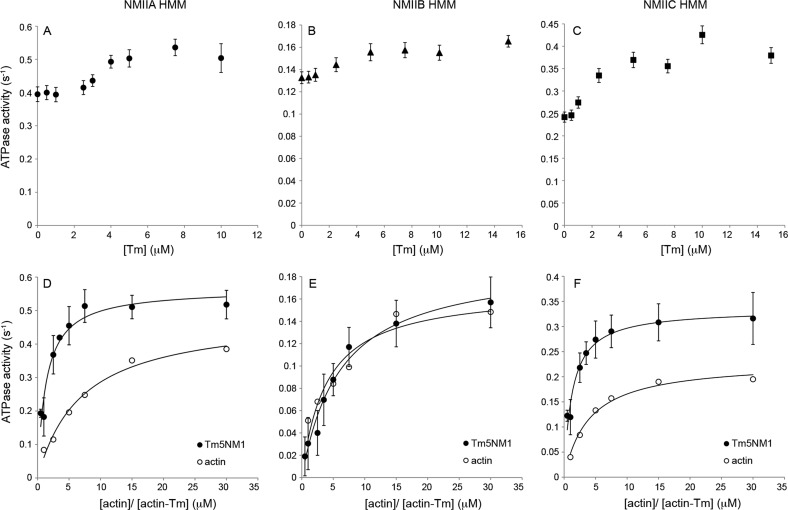
References
-
- Engelhardt W. A.; Ljubimowa M. N. (1939) Myosine and adenosinetriphosphatase. Nature 144, 668–669.
-
- Gordon A. M.; Homsher E.; Regnier M. (2000) Regulation of contraction in striated muscle. Physiol. Rev. 80, 853–924. - PubMed
-
- Bremel R. D.; Weber A. (1972) Cooperation within actin filament in vertebrate skeletal muscle. Nat. New Biol. 238, 97–101. - PubMed
-
- Pruyne D. (2008) Tropomyosin function in yeast. Adv. Exp. Med. Biol. 644, 168–186. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
