

Spatial regulation of Aurora A activity during mitotic spindle assembly requires RHAMM to correctly localize TPX2
- PMID: 24875404
- PMCID: PMC4111680
- DOI: 10.4161/cc.29270
Spatial regulation of Aurora A activity during mitotic spindle assembly requires RHAMM to correctly localize TPX2
Abstract
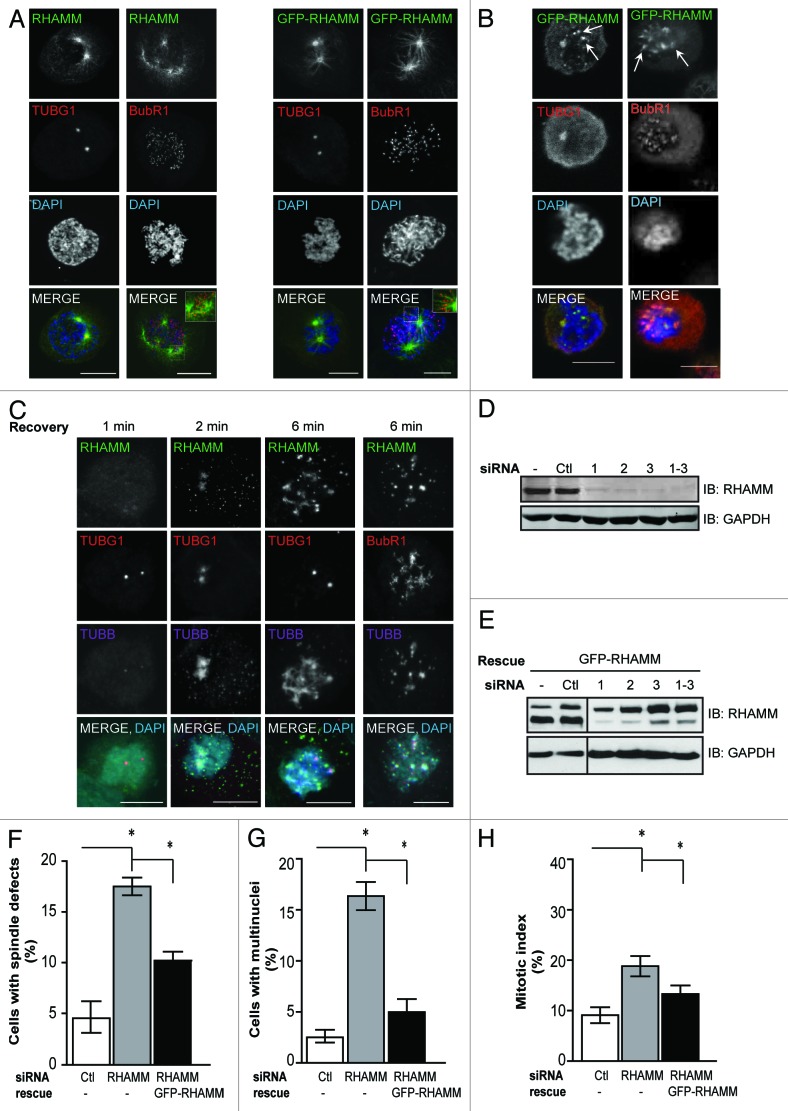
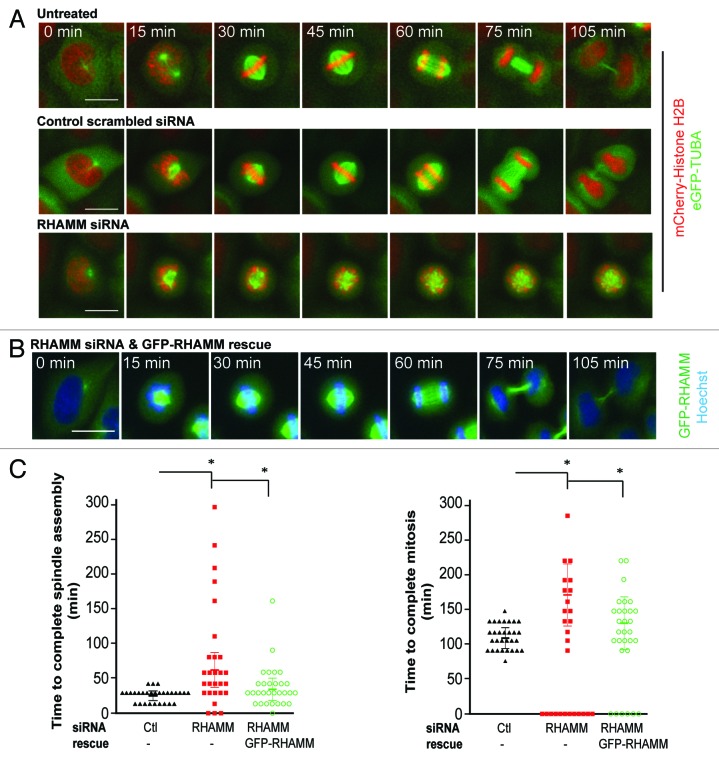
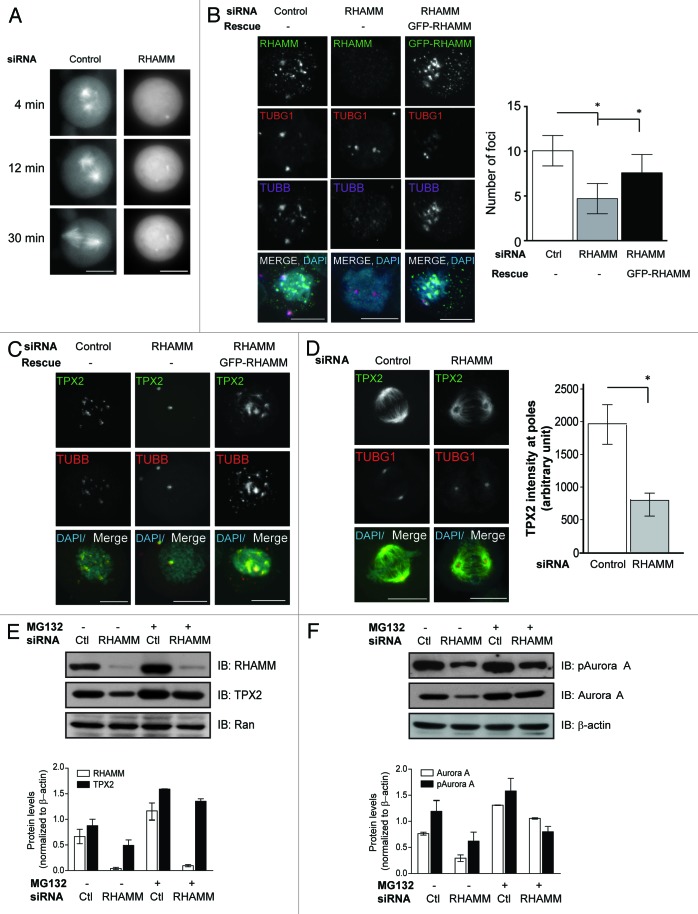
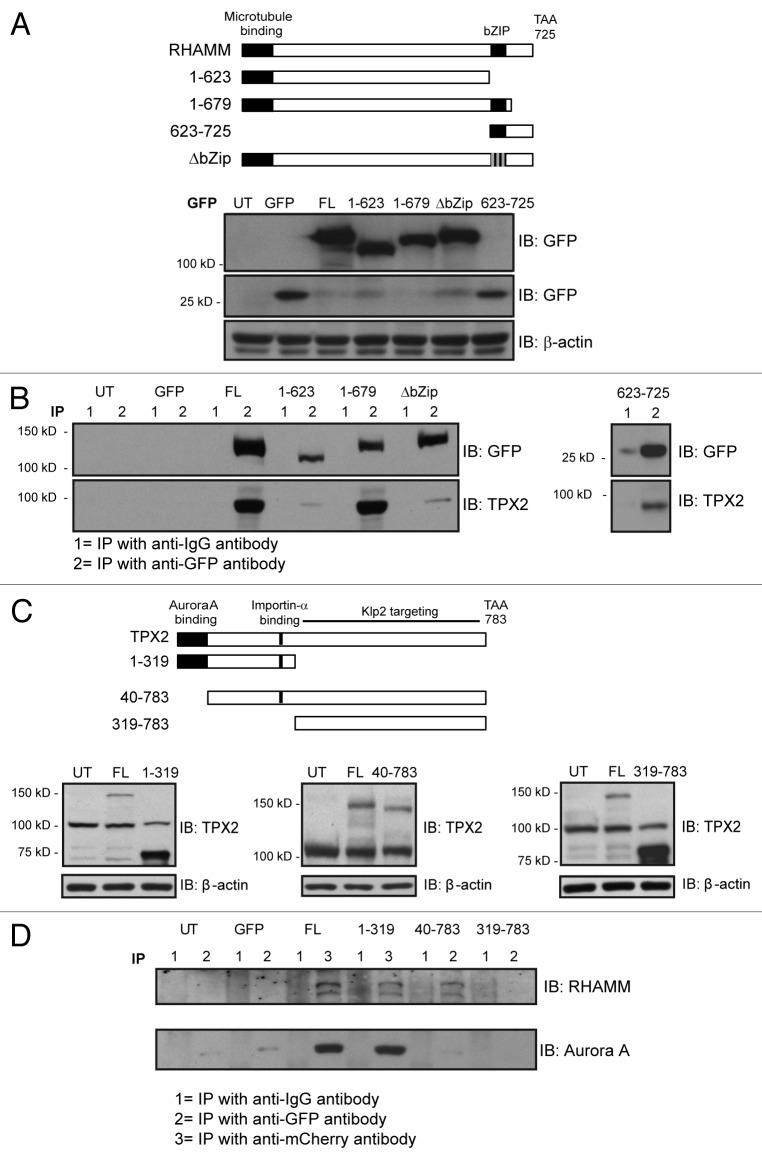
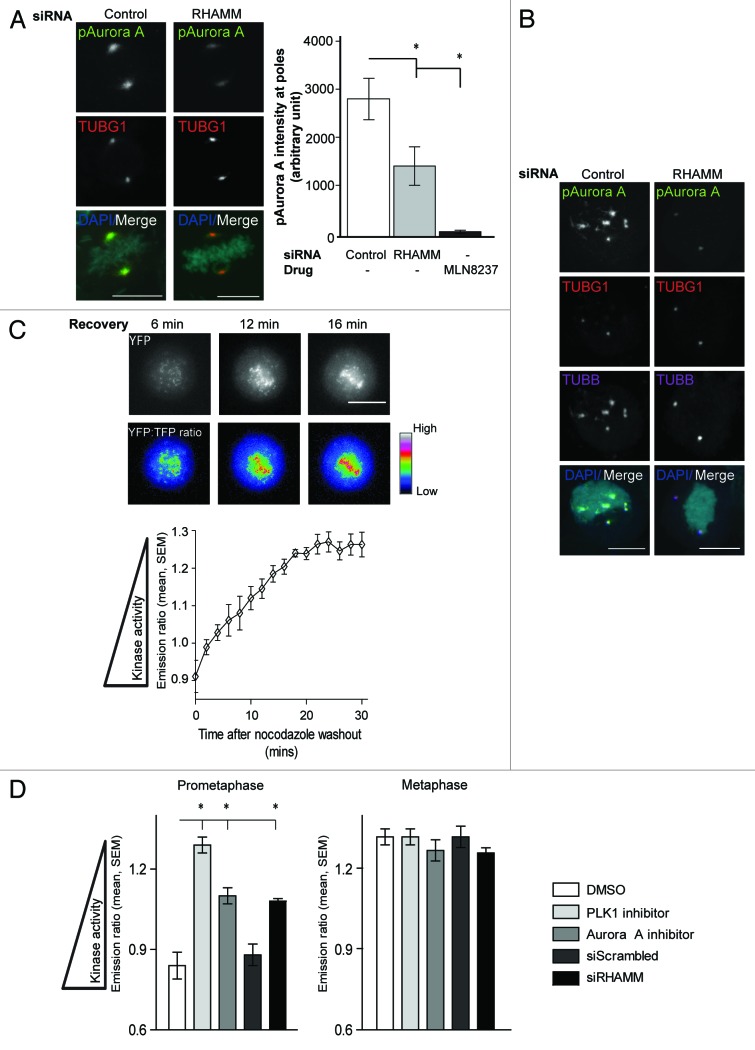
Construction of a mitotic spindle requires biochemical pathways to assemble spindle microtubules and structural proteins to organize these microtubules into a bipolar array. Through a complex with dynein, the receptor for hyaluronan-mediated motility (RHAMM) cross-links mitotic microtubules to provide structural support, maintain spindle integrity, and correctly orient the mitotic spindle. Here, we locate RHAMM to sites of microtubule assembly at centrosomes and non-centrosome sites near kinetochores and demonstrate that RHAMM is required for the activation of Aurora kinase A. Silencing of RHAMM delays the kinetics of spindle assembly, mislocalizes targeting protein for XKlp2 (TPX2), and attenuates the localized activation of Aurora kinase A with a consequent reduction in mitotic spindle length. The RHAMM-TPX2 complex requires a C-terminal basic leucine zipper in RHAMM and a domain that includes the nuclear localization signal in TPX2. Together, our findings identify RHAMM as a critical regulator for Aurora kinase A signaling and suggest that RHAMM ensures bipolar spindle assembly and mitotic progression through the integration of biochemical and structural pathways.
Keywords: Aurora kinase A; RHAMM; TPX2; mitosis; spindle assembly.
Figures

References
-
- Mori D, Yano Y, Toyo-oka K, Yoshida N, Yamada M, Muramatsu M, Zhang D, Saya H, Toyoshima YY, Kinoshita K, et al. NDEL1 phosphorylation by Aurora-A kinase is essential for centrosomal maturation, separation, and TACC3 recruitment. Mol Cell Biol. 2007;27:352–67. doi: 10.1128/MCB.00878-06. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous