

Hypoxia-inducible lipid droplet-associated (HILPDA) is a novel peroxisome proliferator-activated receptor (PPAR) target involved in hepatic triglyceride secretion
- PMID: 24876382
- PMCID: PMC4094041
- DOI: 10.1074/jbc.M114.570044
Hypoxia-inducible lipid droplet-associated (HILPDA) is a novel peroxisome proliferator-activated receptor (PPAR) target involved in hepatic triglyceride secretion
Abstract
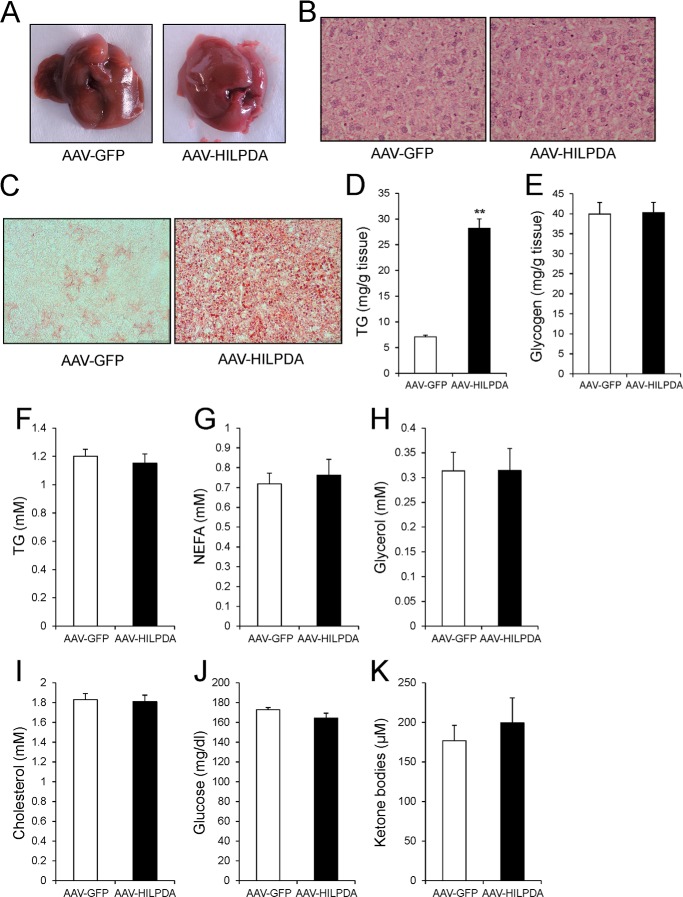
Peroxisome proliferator-activated receptors (PPARs) play major roles in the regulation of hepatic lipid metabolism through the control of numerous genes involved in processes such as lipid uptake and fatty acid oxidation. Here we identify hypoxia-inducible lipid droplet-associated (Hilpda/Hig2) as a novel PPAR target gene and demonstrate its involvement in hepatic lipid metabolism. Microarray analysis revealed that Hilpda is one of the most highly induced genes by the PPARα agonist Wy14643 in mouse precision cut liver slices. Induction of Hilpda mRNA by Wy14643 was confirmed in mouse and human hepatocytes. Oral dosing with Wy14643 similarly induced Hilpda mRNA levels in livers of wild-type mice but not Ppara(-/-) mice. Transactivation studies and chromatin immunoprecipitation showed that Hilpda is a direct PPARα target gene via a conserved PPAR response element located 1200 base pairs upstream of the transcription start site. Hepatic overexpression of HILPDA in mice via adeno-associated virus led to a 4-fold increase in liver triglyceride storage, without any changes in key genes involved in de novo lipogenesis, β-oxidation, or lipolysis. Moreover, intracellular lipase activity was not affected by HILPDA overexpression. Strikingly, HILPDA overexpression significantly impaired hepatic triglyceride secretion. Taken together, our data uncover HILPDA as a novel PPAR target that raises hepatic triglyceride storage via regulation of triglyceride secretion.
Keywords: Hig2; Lipid Droplets; Lipoprotein Secretion; Liver Metabolism; Liver Slices; Nuclear Receptor; PPRE; Peroxisome Proliferator-activated Receptor (PPAR); Steatosis; VLDL.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






Similar articles
-
Hypoxia-inducible lipid droplet-associated induces DGAT1 and promotes lipid storage in hepatocytes.Mol Metab. 2021 May;47:101168. doi: 10.1016/j.molmet.2021.101168. Epub 2021 Jan 16. Mol Metab. 2021. PMID: 33465519 Free PMC article.
-
HILPDA Regulates Lipid Metabolism, Lipid Droplet Abundance, and Response to Microenvironmental Stress in Solid Tumors.Mol Cancer Res. 2019 Oct;17(10):2089-2101. doi: 10.1158/1541-7786.MCR-18-1343. Epub 2019 Jul 15. Mol Cancer Res. 2019. PMID: 31308147 Free PMC article.
-
Hepatic PGC-1beta overexpression induces combined hyperlipidemia and modulates the response to PPARalpha activation.Arterioscler Thromb Vasc Biol. 2007 Dec;27(12):2707-13. doi: 10.1161/ATVBAHA.107.155739. Epub 2007 Oct 11. Arterioscler Thromb Vasc Biol. 2007. PMID: 17932310
-
Regulation of lipid droplet homeostasis by hypoxia inducible lipid droplet associated HILPDA.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Sep;1865(9):158738. doi: 10.1016/j.bbalip.2020.158738. Epub 2020 May 11. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 32417386 Review.
-
The role and regulation of the peroxisome proliferator activated receptor alpha in human liver.Biochimie. 2017 May;136:75-84. doi: 10.1016/j.biochi.2016.12.019. Epub 2017 Jan 8. Biochimie. 2017. PMID: 28077274 Review.
Cited by
-
The Lipid Droplet Protein Hypoxia-inducible Gene 2 Promotes Hepatic Triglyceride Deposition by Inhibiting Lipolysis.J Biol Chem. 2015 Jun 12;290(24):15175-84. doi: 10.1074/jbc.M115.650184. Epub 2015 Apr 28. J Biol Chem. 2015. PMID: 25922078 Free PMC article.
-
Weighted single-step genome-wide association study for direct and maternal genetic effects associated with birth and weaning weights in sheep.Sci Rep. 2024 Jun 7;14(1):13120. doi: 10.1038/s41598-024-63974-0. Sci Rep. 2024. PMID: 38849438 Free PMC article.
-
Hypoxia-Inducible Lipid Droplet-Associated Is Not a Direct Physiological Regulator of Lipolysis in Adipose Tissue.Endocrinology. 2017 May 1;158(5):1231-1251. doi: 10.1210/en.2016-1809. Endocrinology. 2017. PMID: 28323980 Free PMC article.
-
Lipid droplet storage promotes murine pancreatic tumor growth.Oncol Rep. 2021 Apr;45(4):21. doi: 10.3892/or.2021.7972. Epub 2021 Mar 2. Oncol Rep. 2021. PMID: 33649859 Free PMC article.
-
Hypoxia-inducible lipid droplet-associated protein inhibits adipose triglyceride lipase.J Lipid Res. 2018 Mar;59(3):531-541. doi: 10.1194/jlr.M082388. Epub 2018 Jan 11. J Lipid Res. 2018. PMID: 29326160 Free PMC article.
References
-
- McPherson P. A., McEneny J. (2012) The biochemistry of ketogenesis and its role in weight management, neurological disease and oxidative stress. J. Physiol. Biochem. 68, 141–151 - PubMed
-
- Nakamura M. T., Cheon Y., Li Y., Nara T. Y. (2004) Mechanisms of regulation of gene expression by fatty acids. Lipids 39, 1077–1083 - PubMed
-
- Chawla A., Repa J. J., Evans R. M., Mangelsdorf D. J. (2001) Nuclear receptors and lipid physiology: opening the X-files. Science 294, 1866–1870 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
