

Role of 6-O-sulfated heparan sulfate in chronic renal fibrosis
- PMID: 24878958
- PMCID: PMC4106343
- DOI: 10.1074/jbc.M114.554691
Role of 6-O-sulfated heparan sulfate in chronic renal fibrosis
Abstract
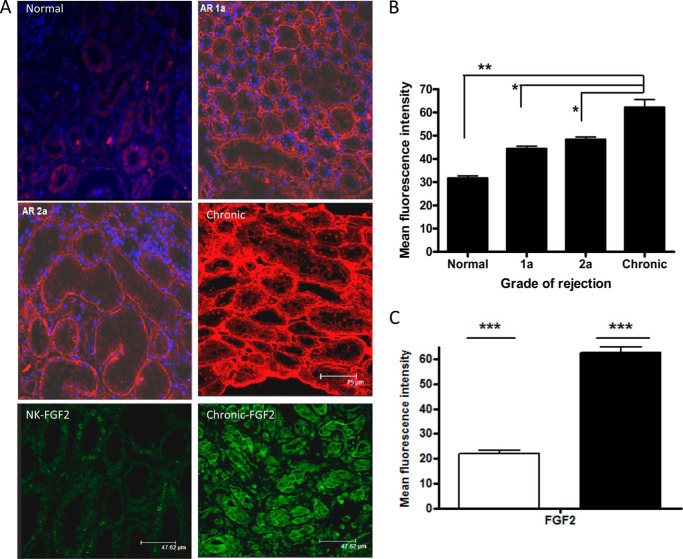
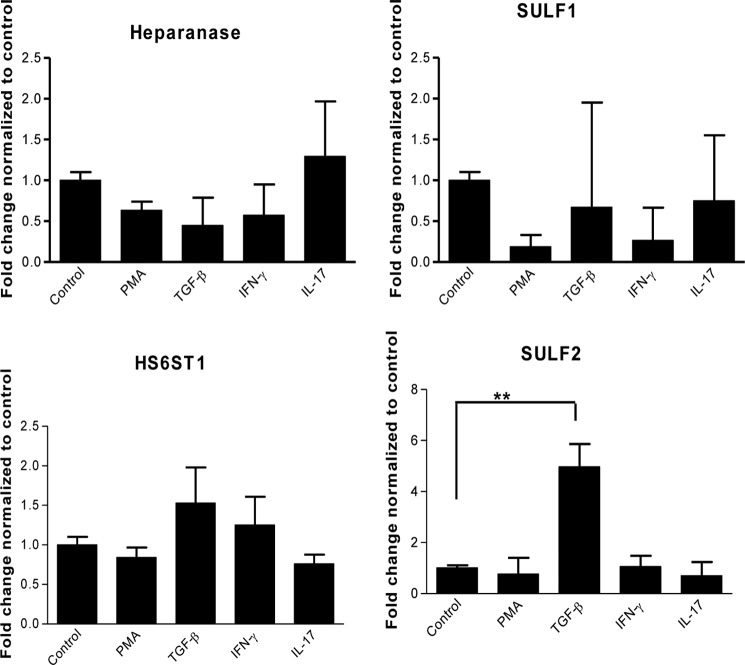
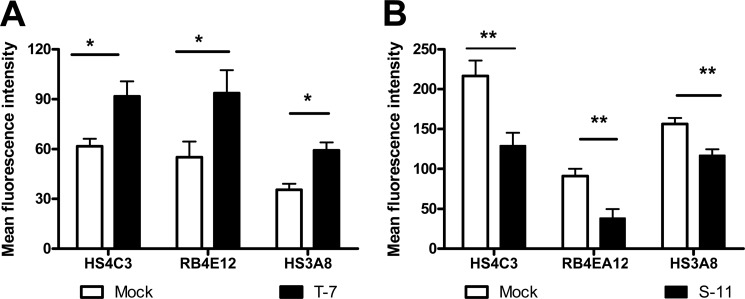
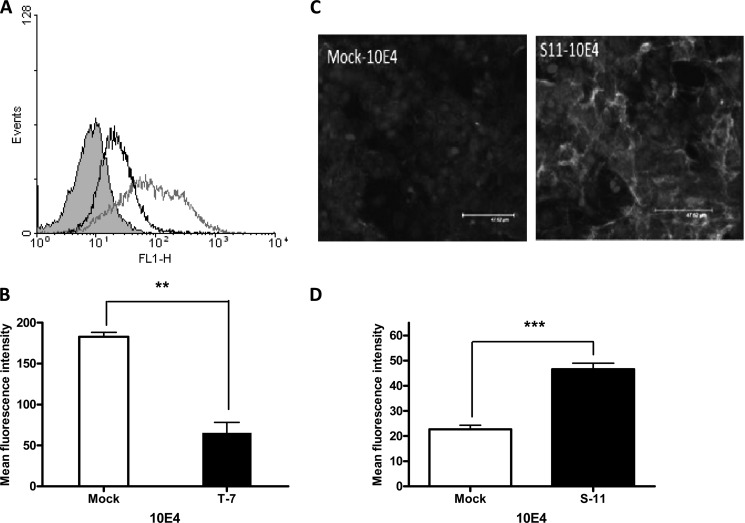
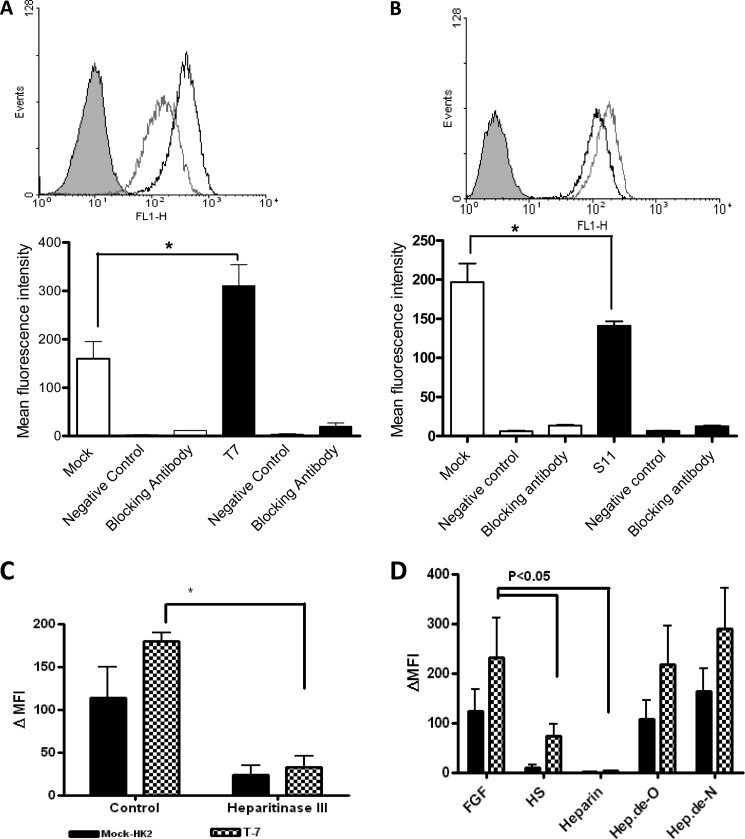
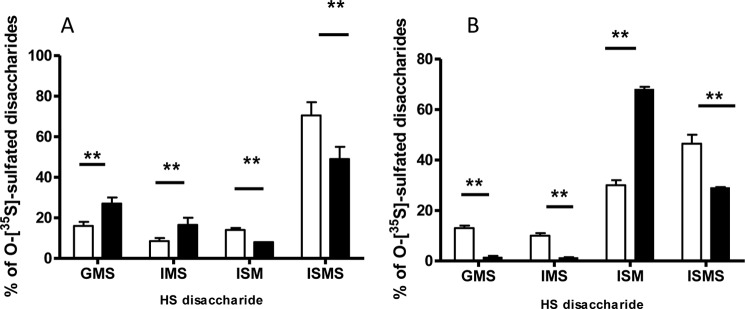
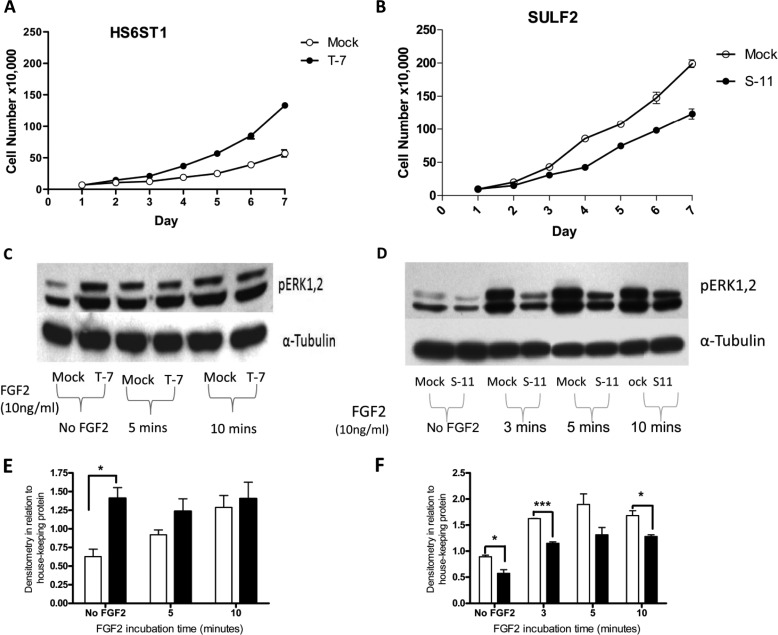
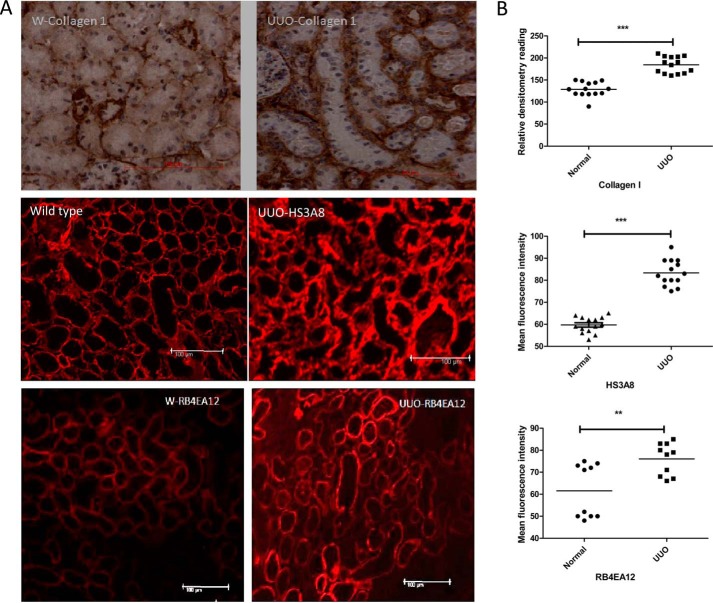
Heparan sulfate (HS) plays a crucial role in the fibrosis associated with chronic allograft dysfunction by binding and presenting cytokines and growth factors to their receptors. These interactions critically depend on the distribution of 6-O-sulfated glucosamine residues, which is generated by glucosaminyl-6-O-sulfotransferases (HS6STs) and selectively removed by cell surface HS-6-O-endosulfatases (SULFs). Using human renal allografts we found increased expression of 6-O-sulfated HS domains in tubular epithelial cells during chronic rejection as compared with the controls. Stimulation of renal epithelial cells with TGF-β induced SULF2 expression. To examine the role of 6-O-sulfated HS in the development of fibrosis, we generated stable HS6ST1 and SULF2 overexpressing renal epithelial cells. Compared with mock transfectants, the HS6ST1 transfectants showed significantly increased binding of FGF2 (p = 0.0086) and pERK activation. HS6ST1 transfectants displayed a relative increase in mono-6-O-sulfated disaccharides accompanied by a decrease in iduronic acid 2-O-sulfated disaccharide structures. In contrast, SULF2 transfectants showed significantly reduced FGF2 binding and phosphorylation of ERK. Structural analysis of HS showed about 40% down-regulation in 6-O-sulfation with a parallel increase in iduronic acid mono-2-O-sulfated disaccharides. To assess the relevance of these data in vivo we established a murine model of fibrosis (unilateral ureteric obstruction (UUO)). HS-specific phage display antibodies (HS3A8 and RB4EA12) showed significant increase in 6-O-sulfation in fibrotic kidney compared with the control. These results suggest an important role of 6-O-sulfation in the pathogenesis of fibrosis associated with chronic rejection.
Keywords: Fibroblast Growth Factor (FGF); Fibrosis; Heparan Sulfate; Sulfotransferase; Transplantation.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Zeisberg M., Neilson E. G. (2010) Mechanisms of tubulointerstitial fibrosis. J. Am. Soc. Nephrol. 21, 1819–1834 - PubMed
-
- Lindahl U., Kusche-Gullberg M., Kjellén L. (1998) Regulated diversity of heparan sulfate. J. Biol. Chem. 273, 24979–24982 - PubMed
-
- Esko J. D., Selleck S. B. (2002) Order out of chaos: assembly of ligand binding sites in heparan sulfate. Annu. Rev. Biochem. 71, 435–471 - PubMed
-
- Feyzi E., Saldeen T., Larsson E., Lindahl U., Salmivirta M. (1998) Age-dependent modulation of heparan sulfate structure and function. J. Biol. Chem. 273, 13395–13398 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
