

The principal role of Ku in telomere length maintenance is promotion of Est1 association with telomeres
- PMID: 24879463
- PMCID: PMC4125388
- DOI: 10.1534/genetics.114.164707
The principal role of Ku in telomere length maintenance is promotion of Est1 association with telomeres
Abstract
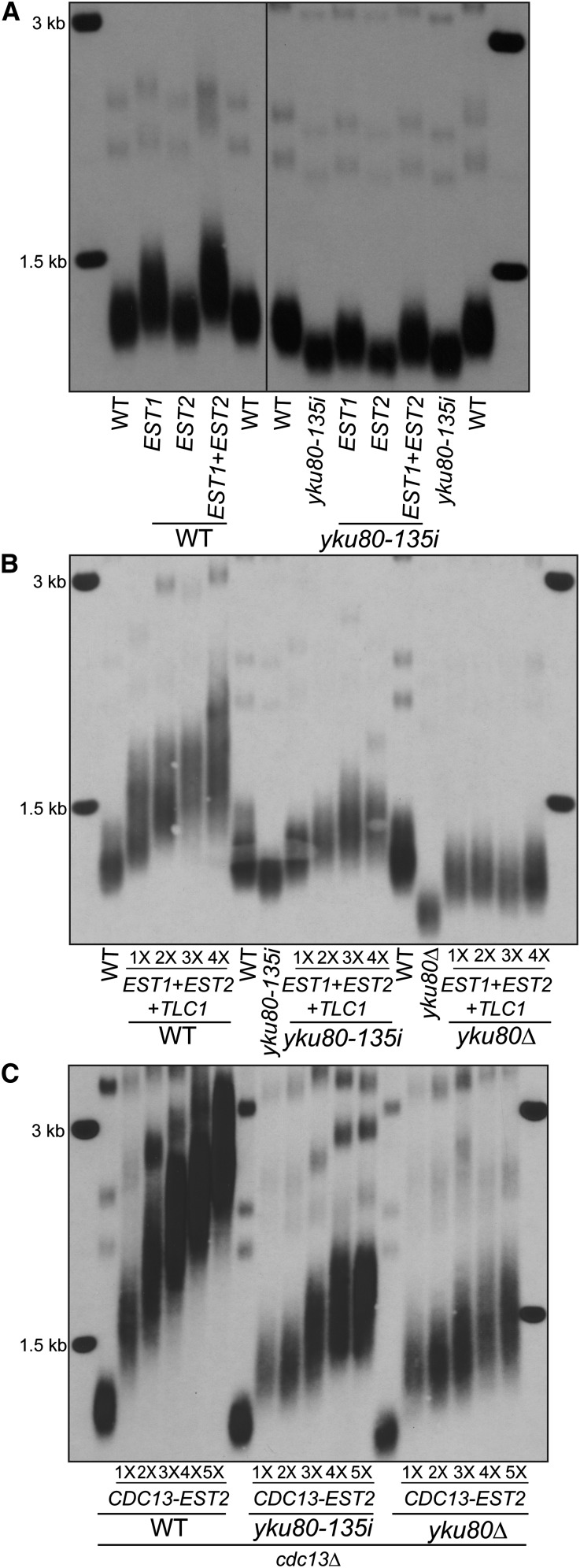
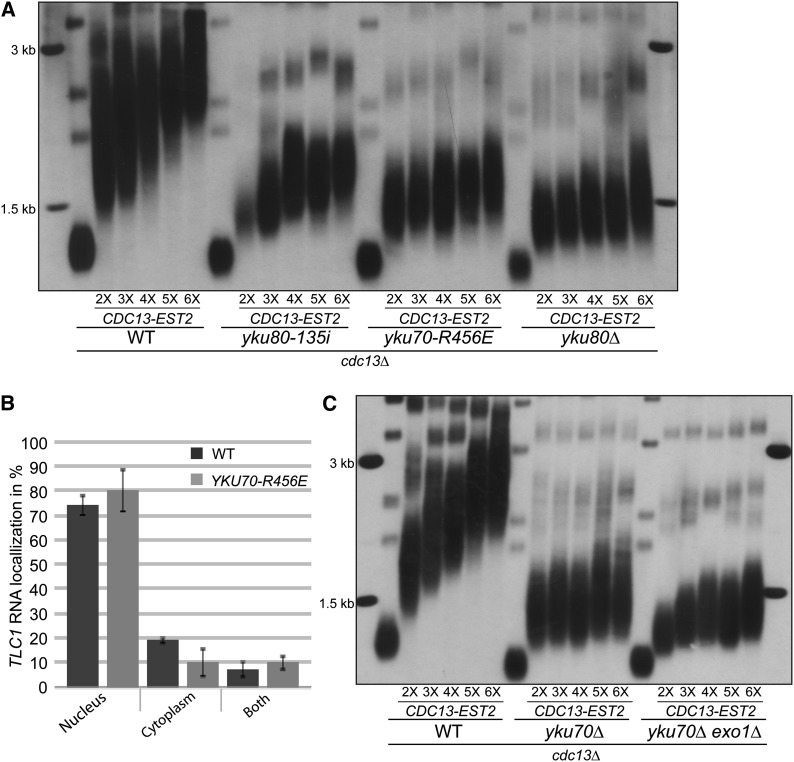
Telomere length is tightly regulated in cells that express telomerase. The Saccharomyces cerevisiae Ku heterodimer, a DNA end-binding complex, positively regulates telomere length in a telomerase-dependent manner. Ku associates with the telomerase RNA subunit TLC1, and this association is required for TLC1 nuclear retention. Ku-TLC1 interaction also impacts the cell-cycle-regulated association of the telomerase catalytic subunit Est2 to telomeres. The promotion of TLC1 nuclear localization and Est2 recruitment have been proposed to be the principal role of Ku in telomere length maintenance, but neither model has been directly tested. Here we study the impact of forced recruitment of Est2 to telomeres on telomere length in the absence of Ku's ability to bind TLC1 or DNA ends. We show that tethering Est2 to telomeres does not promote efficient telomere elongation in the absence of Ku-TLC1 interaction or DNA end binding. Moreover, restoration of TLC1 nuclear localization, even when combined with Est2 recruitment, does not bypass the role of Ku. In contrast, forced recruitment of Est1, which has roles in telomerase recruitment and activation, to telomeres promotes efficient and progressive telomere elongation in the absence of Ku-TLC1 interaction, Ku DNA end binding, or Ku altogether. Ku associates with Est1 and Est2 in a TLC1-dependent manner and enhances Est1 recruitment to telomeres independently of Est2. Together, our results unexpectedly demonstrate that the principal role of Ku in telomere length maintenance is to promote the association of Est1 with telomeres, which may in turn allow for efficient recruitment and activation of the telomerase holoenzyme.
Keywords: Est1; Est2; Ku; TLC1; telomerase; telomere.
Copyright © 2014 by the Genetics Society of America.
Figures





References
-
- Aparicio, O. M., J. V. Geisberg, E. Sekinger, A. Yang, Z. Moqtaderi et al., 2005 Chromatin immunoprecipitation for determining the association of proteins with specific genomic regions in vivo, pp. 21.23.21–21.23.17 in Current Protocols in Molecular Biology, edited by F. M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman et al. John Wiley & Sons, New York. - PubMed
-
- Bianchi A., Negrini S., Shore D., 2004. Delivery of yeast telomerase to a DNA break depends on the recruitment functions of Cdc13 and Est1. Mol. Cell 16: 139–146. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
