

Fungal pathogens are platforms for discovering novel and conserved septin properties
- PMID: 24879478
- PMCID: PMC4266239
- DOI: 10.1016/j.mib.2014.04.004
Fungal pathogens are platforms for discovering novel and conserved septin properties
Abstract
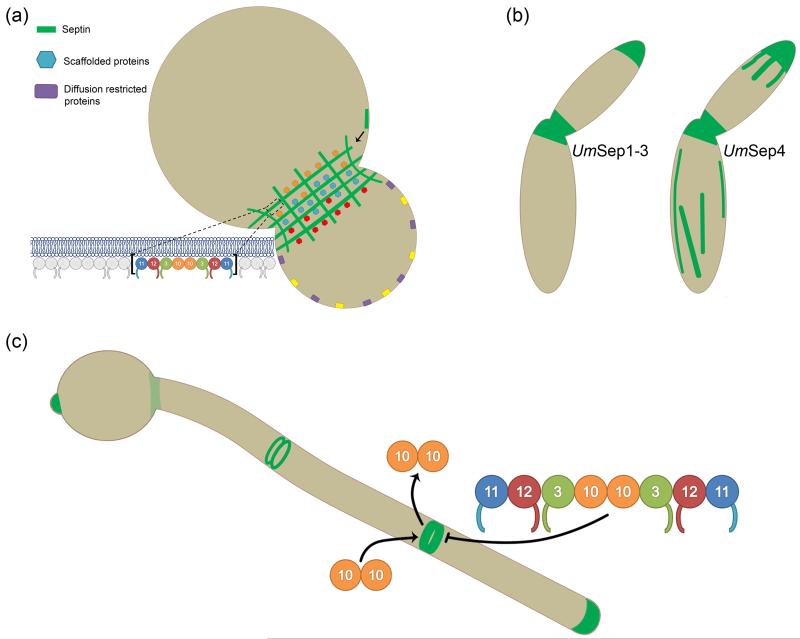
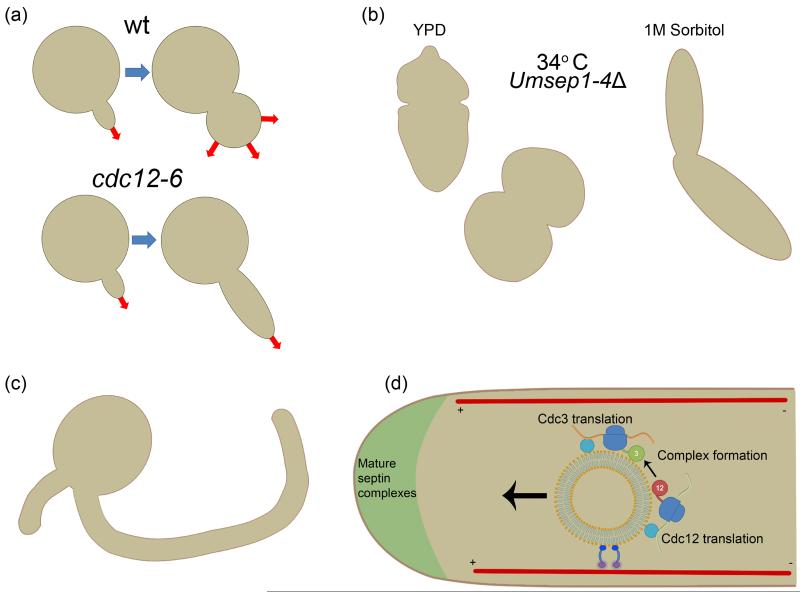
Septins are filament-forming GTP-binding proteins that act as scaffolds in diverse cell functions including division, polarity and membrane remodeling. In a variety of fungal pathogens, it has been observed that septins are required for virulence because cells are unable to survive or are misshapen when septins are mutated. Cell morphology is interconnected with pathogenesis and thus septin mutants displaying aberrant cell morphologies are commonly deficient in host tissue invasion. The degree to which septins orchestrate versus maintain changes in fungal cell morphology during pathogenesis remains to be determined. Aside from the importance of septins in the process of pathogenesis, animal and plant fungal pathogens display complexity in septin form, dynamics, and function not seen in Saccharomyces cerevisiae making these organisms important models for uncovering diversity in septin behavior. Additionally, host septins have recently been implicated in the process of Candida albicans invasion, motivating the need to examine host septins in fungal pathogenesis. Understanding the role of septins in the host-pathogen interaction not only illuminates pathogenesis mechanisms but importantly also expands our understanding of septin biology in general.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Phosphoregulation of Nap1 plays a role in septin ring dynamics and morphogenesis in Candida albicans.mBio. 2014 Feb 4;5(1):e00915-13. doi: 10.1128/mBio.00915-13. mBio. 2014. PMID: 24496790 Free PMC article.
-
Insights of roles played by septins in pathogenic fungi.Virulence. 2021 Dec;12(1):1550-1562. doi: 10.1080/21505594.2021.1933370. Virulence. 2021. PMID: 34097566 Free PMC article. Review.
-
Rsr1 Palmitoylation and GTPase Activity Status Differentially Coordinate Nuclear, Septin, and Vacuole Dynamics in Candida albicans.mBio. 2020 Oct 13;11(5):e01666-20. doi: 10.1128/mBio.01666-20. mBio. 2020. PMID: 33051364 Free PMC article.
-
Candida albicans septin mutants are defective for invasive growth and virulence.Infect Immun. 2003 Jul;71(7):4045-51. doi: 10.1128/IAI.71.7.4045-4051.2003. Infect Immun. 2003. PMID: 12819094 Free PMC article.
-
Forging the ring: from fungal septins' divergent roles in morphology, septation and virulence to factors contributing to their assembly into higher order structures.Microbiology (Reading). 2016 Sep;162(9):1527-1534. doi: 10.1099/mic.0.000359. Epub 2016 Aug 23. Microbiology (Reading). 2016. PMID: 27559018 Free PMC article. Review.
Cited by
-
Discovery of broad-spectrum fungicides that block septin-dependent infection processes of pathogenic fungi.Nat Microbiol. 2020 Dec;5(12):1565-1575. doi: 10.1038/s41564-020-00790-y. Epub 2020 Sep 21. Nat Microbiol. 2020. PMID: 32958858
-
Septin 9 induces lipid droplets growth by a phosphatidylinositol-5-phosphate and microtubule-dependent mechanism hijacked by HCV.Nat Commun. 2016 Jul 15;7:12203. doi: 10.1038/ncomms12203. Nat Commun. 2016. PMID: 27417143 Free PMC article.
-
MCC/Eisosomes Regulate Cell Wall Synthesis and Stress Responses in Fungi.J Fungi (Basel). 2017 Nov 3;3(4):61. doi: 10.3390/jof3040061. J Fungi (Basel). 2017. PMID: 29371577 Free PMC article. Review.
-
Mechanisms of Cytokinesis in Basidiomycetous Yeasts.Fungal Biol Rev. 2017 Mar;31(2):73-87. doi: 10.1016/j.fbr.2016.12.002. Epub 2017 Jan 12. Fungal Biol Rev. 2017. PMID: 28943887 Free PMC article.
-
Expression of Septin4 in Schistosoma japonicum-infected mouse livers after praziquantel treatment.Parasit Vectors. 2015 Jan 13;8:19. doi: 10.1186/s13071-015-0640-9. Parasit Vectors. 2015. PMID: 25582427 Free PMC article.
References
-
- Beise N, Trimble W. Septins at a glance. J Cell Sci. 2011;124:4141–4146. - PubMed
-
- Mostowy S, Cossart P. Septins: the fourth component of the cytoskeleton. Nat Rev Mol Cell Biol. 2012;13:183–194. - PubMed
-
- Gladfelter AS, Pringle JR, Lew DJ. The septin cortex at the yeast mother-bud neck. Curr Opin Microbiol. 2001;4:681–689. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
