

FHOD1 interaction with nesprin-2G mediates TAN line formation and nuclear movement
- PMID: 24880667
- PMCID: PMC4113092
- DOI: 10.1038/ncb2981
FHOD1 interaction with nesprin-2G mediates TAN line formation and nuclear movement
Abstract
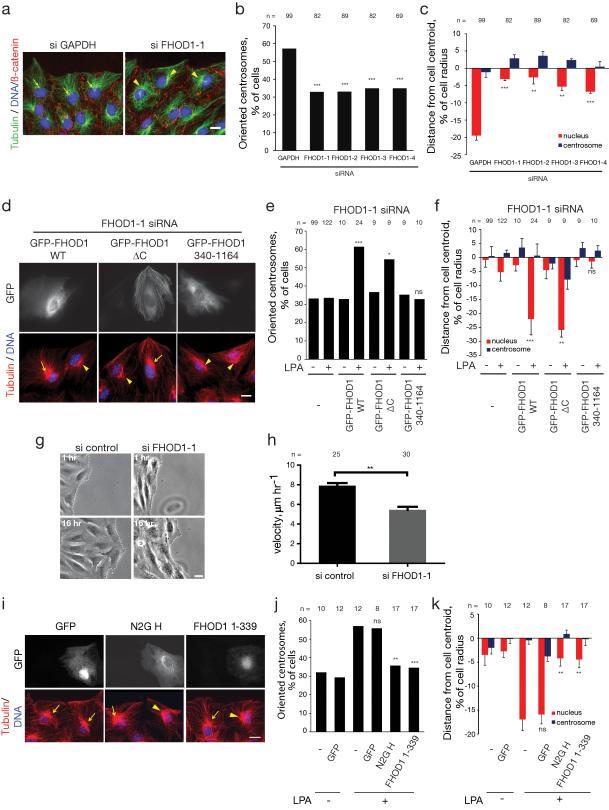
Active positioning of the nucleus is integral to division, migration and differentiation of mammalian cells. Fibroblasts polarizing for migration orient their centrosomes by actin-dependent nuclear movement. This nuclear movement depends on nesprin-2 giant (N2G), a large, actin-binding outer nuclear membrane component of transmembrane actin-associated (TAN) lines that couple nuclei to moving actin cables. Here, we identify the diaphanous formin FHOD1 as an interaction partner of N2G. Silencing FHOD1 expression or expression of fragments containing binding sites for N2G or FHOD1 disrupted nuclear movement and centrosome orientation in polarizing fibroblasts. Unexpectedly, silencing of FHOD1 expression did not affect the formation or rearward flow of dorsal actin cables required for nuclear positioning. Rather, N2G-FHOD1 interaction provided a second connection to actin cables essential for TAN line formation and thus nuclear movement. These results reveal a unique function for a formin in coupling an organelle to actin filaments for translocation, and suggest that TAN lines require multi-point attachments to actin cables to resist the large forces necessary to move the nucleus.
Figures




Similar articles
-
Reinforcing the LINC complex connection to actin filaments: the role of FHOD1 in TAN line formation and nuclear movement.Cell Cycle. 2015;14(14):2200-5. doi: 10.1080/15384101.2015.1053665. Epub 2015 Jun 17. Cell Cycle. 2015. PMID: 26083340 Free PMC article.
-
TorsinA controls TAN line assembly and the retrograde flow of dorsal perinuclear actin cables during rearward nuclear movement.J Cell Biol. 2017 Mar 6;216(3):657-674. doi: 10.1083/jcb.201507113. Epub 2017 Feb 27. J Cell Biol. 2017. PMID: 28242745 Free PMC article.
-
Linker of nucleoskeleton and cytoskeleton (LINC) complex-mediated actin-dependent nuclear positioning orients centrosomes in migrating myoblasts.Nucleus. 2015;6(1):77-88. doi: 10.1080/19491034.2015.1004947. Nucleus. 2015. PMID: 25587885 Free PMC article.
-
Formin-binding proteins: modulators of formin-dependent actin polymerization.Biochim Biophys Acta. 2010 Feb;1803(2):174-82. doi: 10.1016/j.bbamcr.2009.06.002. Epub 2009 Jul 7. Biochim Biophys Acta. 2010. PMID: 19589360 Review.
-
FHODs: Nuclear tethered formins for nuclear mechanotransduction.Front Cell Dev Biol. 2023 May 4;11:1160219. doi: 10.3389/fcell.2023.1160219. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37215084 Free PMC article. Review.
Cited by
-
Chain reaction: LINC complexes and nuclear positioning.F1000Res. 2019 Jan 31;8:F1000 Faculty Rev-136. doi: 10.12688/f1000research.16877.1. eCollection 2019. F1000Res. 2019. PMID: 30774932 Free PMC article. Review.
-
Centrifugal Displacement of Nuclei Reveals Multiple LINC Complex Mechanisms for Homeostatic Nuclear Positioning.Curr Biol. 2017 Oct 23;27(20):3097-3110.e5. doi: 10.1016/j.cub.2017.08.073. Epub 2017 Oct 5. Curr Biol. 2017. PMID: 28988861 Free PMC article.
-
Fascin Regulates Nuclear Movement and Deformation in Migrating Cells.Dev Cell. 2016 Aug 22;38(4):371-83. doi: 10.1016/j.devcel.2016.07.021. Dev Cell. 2016. PMID: 27554857 Free PMC article.
-
LINC complex protein nesprin-2 has pro-apoptotic activity via Bcl-2 family proteins.Cell Death Discov. 2024 Jan 15;10(1):29. doi: 10.1038/s41420-023-01763-w. Cell Death Discov. 2024. PMID: 38225256 Free PMC article.
-
Nuclear migration events throughout development.J Cell Sci. 2016 May 15;129(10):1951-61. doi: 10.1242/jcs.179788. J Cell Sci. 2016. PMID: 27182060 Free PMC article. Review.
References
-
- Gomes ER, Jani S, Gundersen GG. Nuclear movement regulated by Cdc42, MRCK, myosin, and actin flow establishes MTOC polarization in migrating cells. Cell. 2005;121:451–463. - PubMed
-
- Chesarone MA, DuPage AG, Goode BL. Unleashing formins to remodel the actin and microtubule cytoskeletons. Nat. Rev. Mol. Cell Biol. 2010;11:62–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
