

The physiology of the pedunculopontine nucleus: implications for deep brain stimulation
- PMID: 24880787
- PMCID: PMC4484763
- DOI: 10.1007/s00702-014-1243-x
The physiology of the pedunculopontine nucleus: implications for deep brain stimulation
Abstract
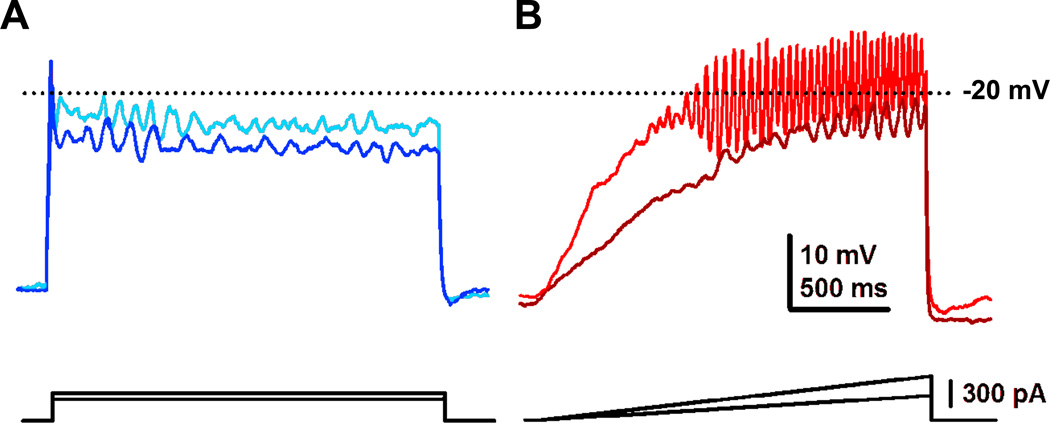
This brief review resolves a number of persistent conflicts regarding the location and characteristics of the mesencephalic locomotor region, which has in the past been described as not locomotion-specific and is more likely the pedunculopontine nucleus (PPN). The parameters of stimulation used to elicit changes in posture and locomotion we now know are ideally suited to match the intrinsic membrane properties of PPN neurons. The physiology of these cells is important not only because it is a major element of the reticular activating system, but also because it is a novel target for the treatment of gait and postural deficits in Parkinson's disease (PD). The discussion explains many of the effects reported following deep brain stimulation (DBS) of the PPN by different groups and provides guidelines for the determination of long-term assessment and effects of PPN DBS. A greater understanding of the physiology of the target nuclei within the brainstem and basal ganglia, amassed over the past decades, has enabled increasingly better patient outcomes from DBS for movement disorders. Despite these improvements, there remains a great opportunity for further understanding of the mechanisms through which DBS has its effects and for further development of appropriate technology to effect these treatments. We review the scientific basis for one of the newest targets, the PPN, in the treatment of PD and other movement disorders, and address the needs for further investigation.
Conflict of interest statement
Figures



References
-
- Alam M, Schwabe K, Krauss JK. The pedunculopontine nucleus area: critical evaluation of interspecies differences relevant for its use as a target for deep brain stimulation. Brain. 2011;134:11–23. - PubMed
-
- Alessandro S, Ceravolo R, Brusa L, Pierantozzi M, Costa A, Galati S, Placidi F, Romigi A, Iani C, Marzetti F, Peppe A. Non-motor functions in parkinsonian patients implamted in the pedunculopontine nucleus: Focus on sleep and cognitive problems. J Neurol Sci. 2010;289:44–48. - PubMed
-
- Aviles-Olmos I, Foltynie T, Panicker J, Cowie D, Limousin P, Hariz M, Fowler CJ, Zrinzo L. Urinary incontinence following deep brain stimulation of the pedunculopontine nucleus. Acta Neurochir (Wien) 2011;153(12):2357–2360. - PubMed
-
- Baghdoyan HA, Rodrigo-Angulo ML, McCarley RW, Hobson JA. Site-specific enhancement and suppression of desynchronized sleep signs following cholinergic stimulation of three brainstem regions. Brain Res. 1984;306:39–52. - PubMed
-
- Chase MH, Morales FR. The control of motoneurons during sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practice of Sleep Medicine. London: WB Saunders; 1994. pp. 163–176.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
