

A presynaptic role for PKA in synaptic tagging and memory
- PMID: 24882624
- PMCID: PMC4143446
- DOI: 10.1016/j.nlm.2014.05.005
A presynaptic role for PKA in synaptic tagging and memory
Abstract
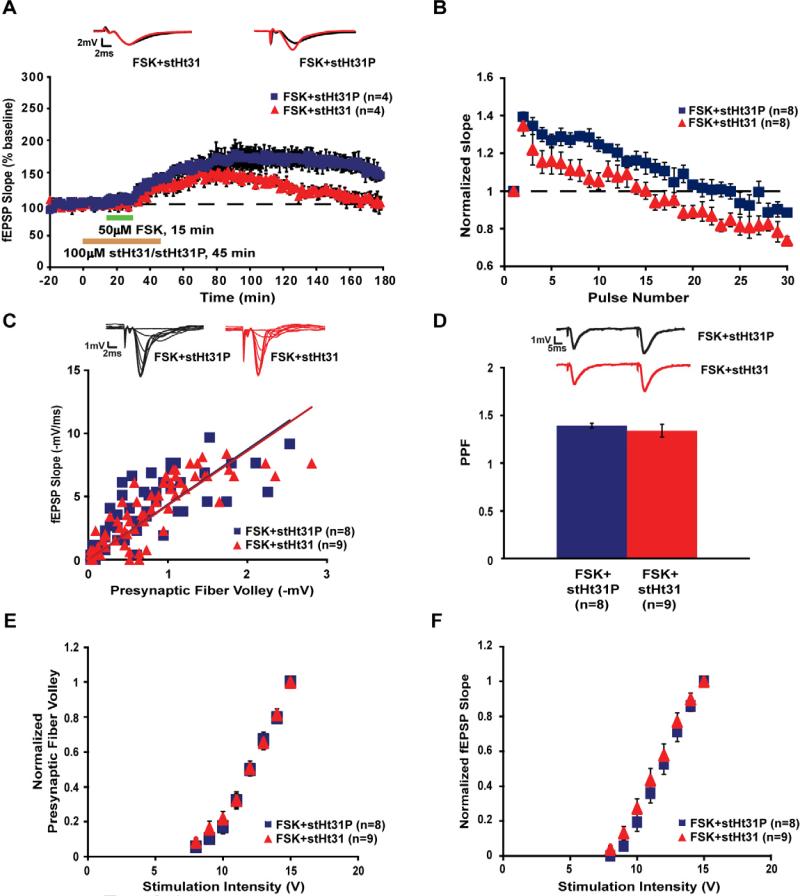
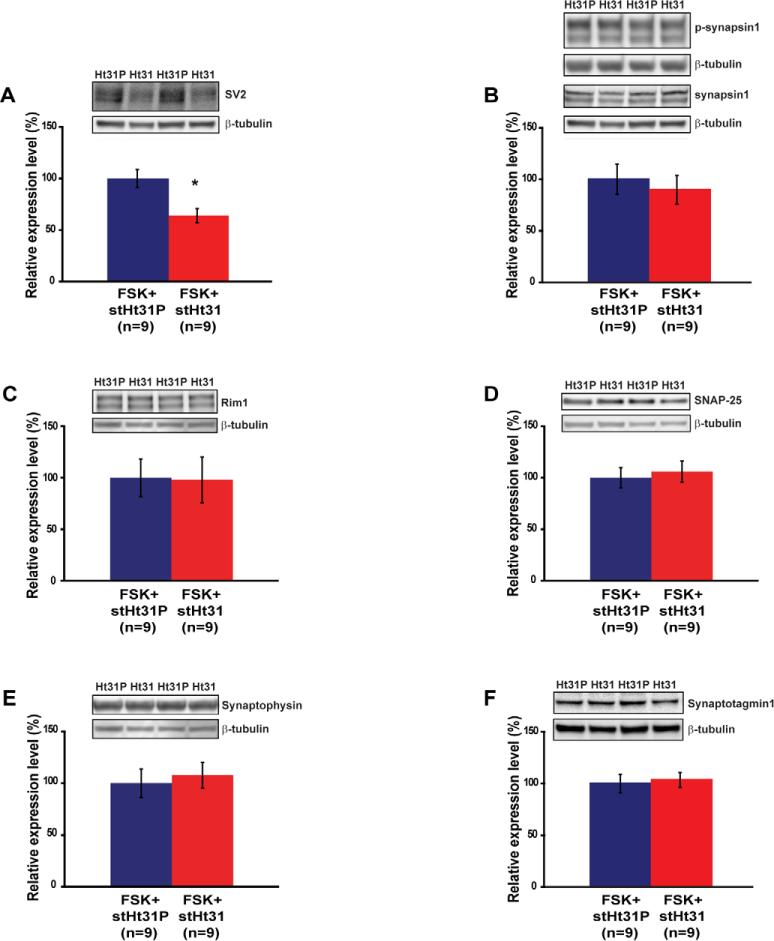
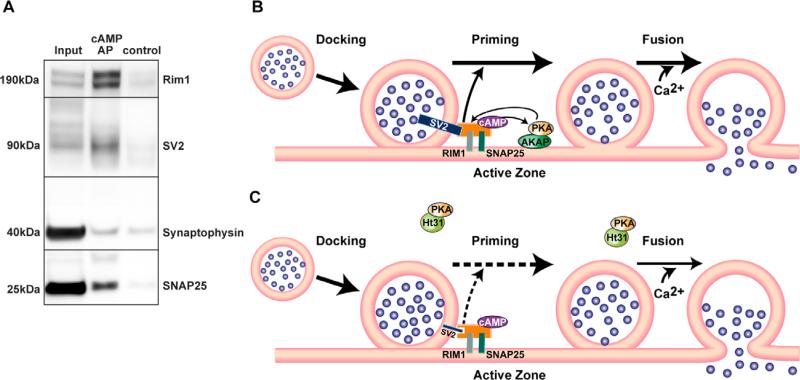
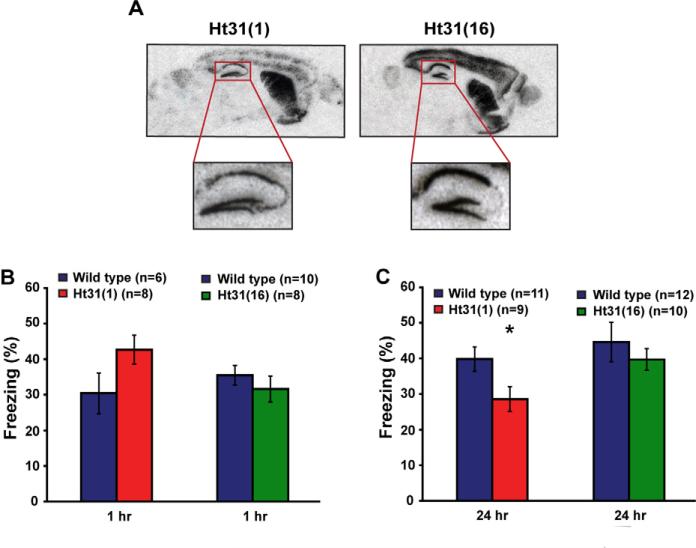
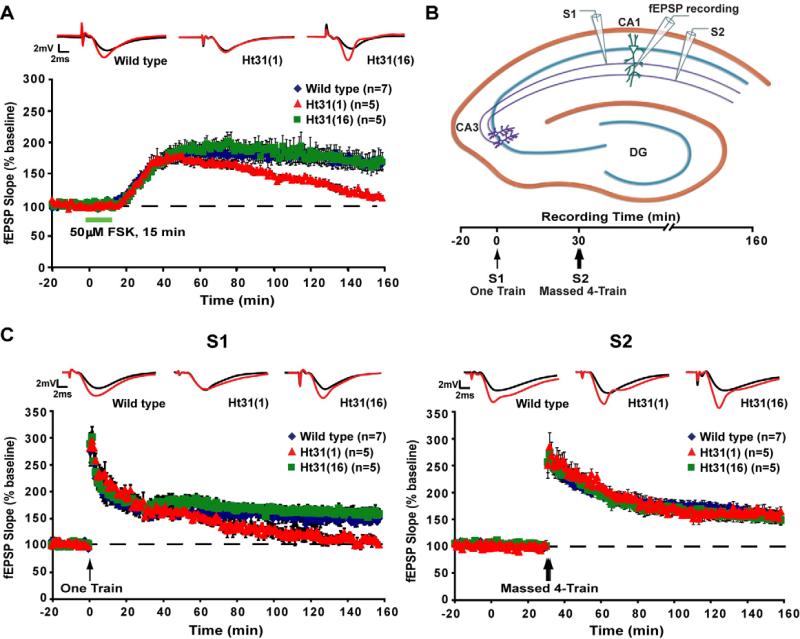
Protein kinase A (PKA) and other signaling molecules are spatially restricted within neurons by A-kinase anchoring proteins (AKAPs). Although studies on compartmentalized PKA signaling have focused on postsynaptic mechanisms, presynaptically anchored PKA may contribute to synaptic plasticity and memory because PKA also regulates presynaptic transmitter release. Here, we examine this issue using genetic and pharmacological application of Ht31, a PKA anchoring disrupting peptide. At the hippocampal Schaffer collateral CA3-CA1 synapse, Ht31 treatment elicits a rapid decay of synaptic responses to repetitive stimuli, indicating a fast depletion of the readily releasable pool of synaptic vesicles. The interaction between PKA and proteins involved in producing this pool of synaptic vesicles is supported by biochemical assays showing that synaptic vesicle protein 2 (SV2), Rim1, and SNAP25 are components of a complex that interacts with cAMP. Moreover, acute treatment with Ht31 reduces the levels of SV2. Finally, experiments with transgenic mouse lines, which express Ht31 in excitatory neurons at the Schaffer collateral CA3-CA1 synapse, highlight a requirement for presynaptically anchored PKA in pathway-specific synaptic tagging and long-term contextual fear memory. These results suggest that a presynaptically compartmentalized PKA is critical for synaptic plasticity and memory by regulating the readily releasable pool of synaptic vesicles.
Keywords: Ht31; PKA; PKA anchoring; SV2; Synaptic tagging.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
SAD-B kinase regulates pre-synaptic vesicular dynamics at hippocampal Schaffer collateral synapses and affects contextual fear memory.J Neurochem. 2016 Jan;136(1):36-47. doi: 10.1111/jnc.13379. Epub 2015 Nov 6. J Neurochem. 2016. PMID: 26444684
-
Inhibition of PKA anchoring to A-kinase anchoring proteins impairs consolidation and facilitates extinction of contextual fear memories.Neurobiol Learn Mem. 2008 Jul;90(1):223-9. doi: 10.1016/j.nlm.2008.03.008. Epub 2008 Apr 28. Neurobiol Learn Mem. 2008. PMID: 18442935
-
Genetic disruption of protein kinase A anchoring reveals a role for compartmentalized kinase signaling in theta-burst long-term potentiation and spatial memory.J Neurosci. 2007 Sep 19;27(38):10278-88. doi: 10.1523/JNEUROSCI.1602-07.2007. J Neurosci. 2007. PMID: 17881534 Free PMC article.
-
Regulation of neuronal PKA signaling through AKAP targeting dynamics.Eur J Cell Biol. 2006 Jul;85(7):627-33. doi: 10.1016/j.ejcb.2006.01.010. Epub 2006 Feb 28. Eur J Cell Biol. 2006. PMID: 16504338 Review.
-
Modulation of hippocampal synaptic transmission and plasticity by neurotrophins.Prog Brain Res. 2000;128:231-41. doi: 10.1016/S0079-6123(00)28020-5. Prog Brain Res. 2000. PMID: 11105682 Review. No abstract available.
Cited by
-
Clocking In Time to Gate Memory Processes: The Circadian Clock Is Part of the Ins and Outs of Memory.Neural Plast. 2018 Apr 12;2018:6238989. doi: 10.1155/2018/6238989. eCollection 2018. Neural Plast. 2018. PMID: 29849561 Free PMC article. Review.
-
Pharmacological Activators of the NR4A Nuclear Receptors Enhance LTP in a CREB/CBP-Dependent Manner.Neuropsychopharmacology. 2017 May;42(6):1243-1253. doi: 10.1038/npp.2016.253. Epub 2016 Nov 11. Neuropsychopharmacology. 2017. PMID: 27834392 Free PMC article.
-
Activation of α7 nicotinic acetylcholine receptors increases intracellular cAMP levels via activation of AC1 in hippocampal neurons.Neuropharmacology. 2015 Aug;95:405-14. doi: 10.1016/j.neuropharm.2015.04.016. Epub 2015 Apr 29. Neuropharmacology. 2015. PMID: 25937212 Free PMC article.
-
Acute Ethanol Inhibition of γ Oscillations Is Mediated by Akt and GSK3β.Front Cell Neurosci. 2016 Aug 17;10:189. doi: 10.3389/fncel.2016.00189. eCollection 2016. Front Cell Neurosci. 2016. PMID: 27582689 Free PMC article.
-
A role for autophagy in long-term spatial memory formation in male rodents.J Neurosci Res. 2018 Mar;96(3):416-426. doi: 10.1002/jnr.24121. Epub 2017 Dec 12. J Neurosci Res. 2018. PMID: 29230855 Free PMC article.
References
-
- Ahmari SE, Buchanan J, Smith SJ. Assembly of presynaptic active zones from cytoplasmic transport packets. Nature Neuroscience. 2000;3(5):445–51. - PubMed
-
- Barco A, Alarcon JM, Kandel ER. Expression of constitutively active CREB protein facilitates the late phase of long-term potentiation by enhancing synaptic capture. Cell. 2002;108(5):689–703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous