

Infection mobilizes hematopoietic stem cells through cooperative NOD-like receptor and Toll-like receptor signaling
- PMID: 24882704
- PMCID: PMC4085166
- DOI: 10.1016/j.chom.2014.05.004
Infection mobilizes hematopoietic stem cells through cooperative NOD-like receptor and Toll-like receptor signaling
Abstract
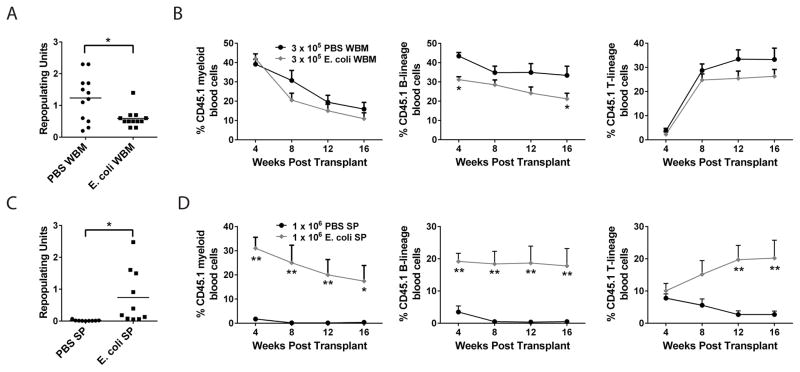
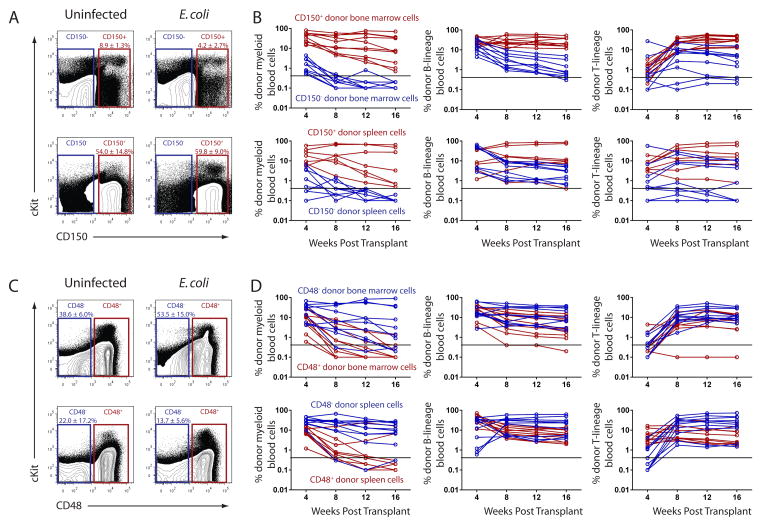
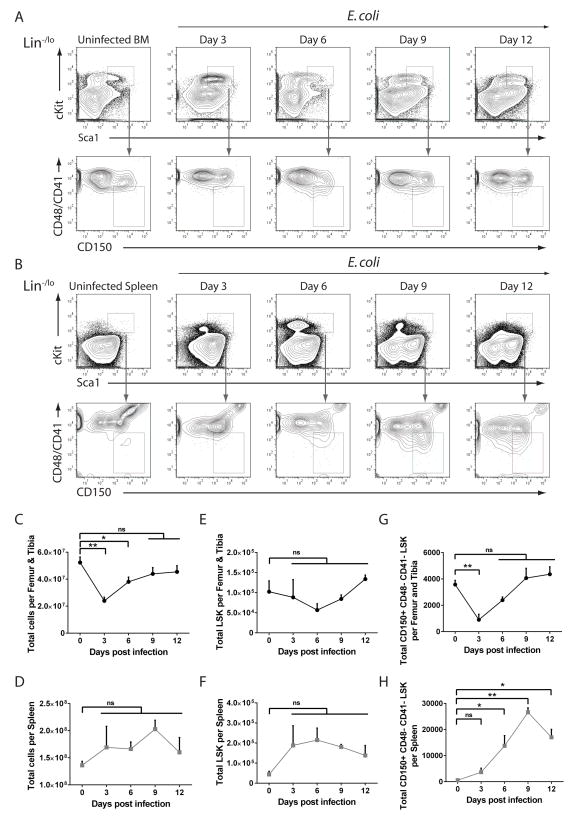
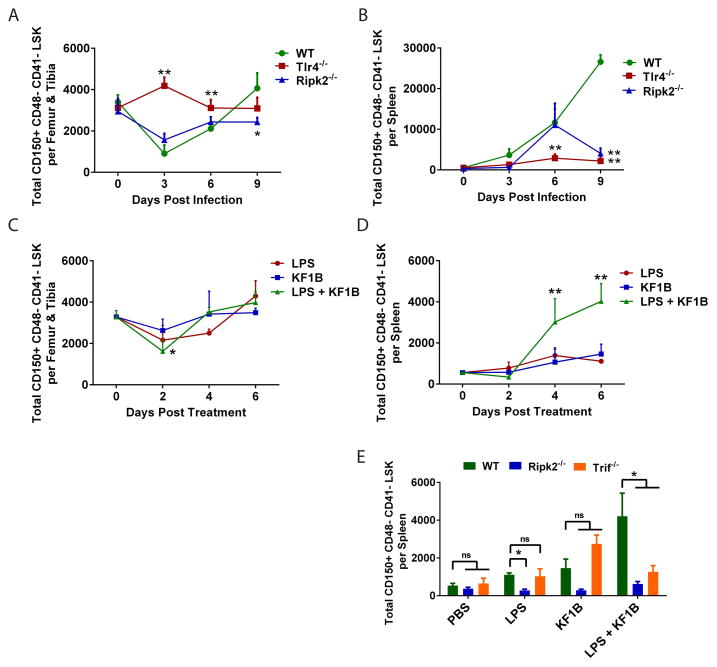
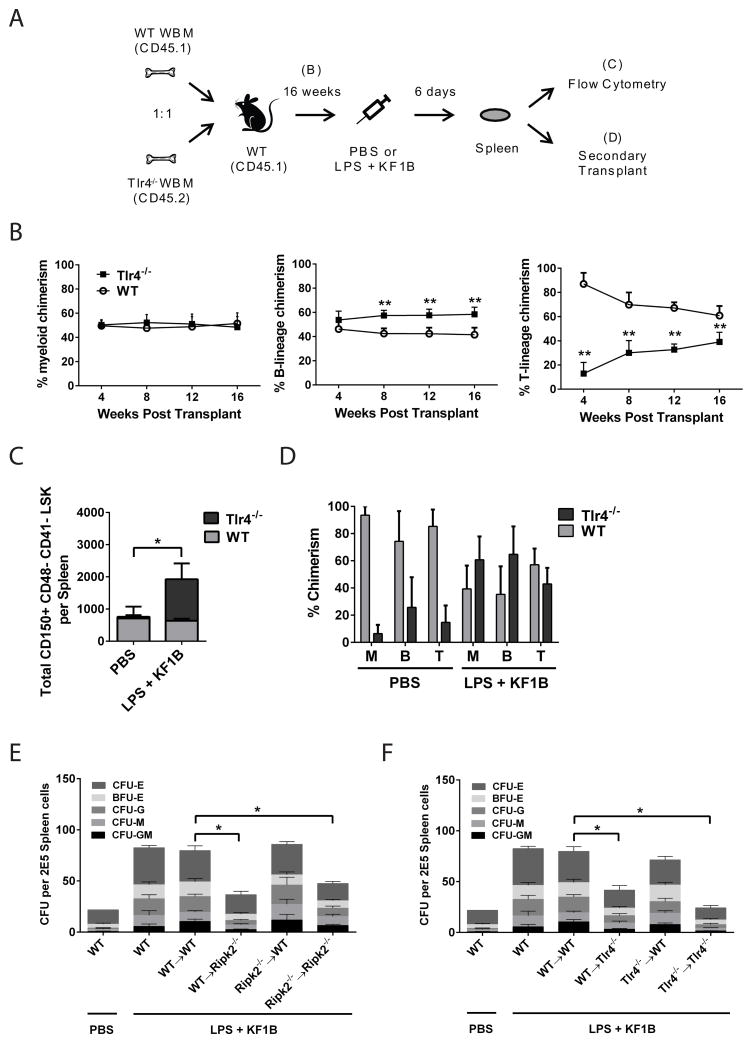
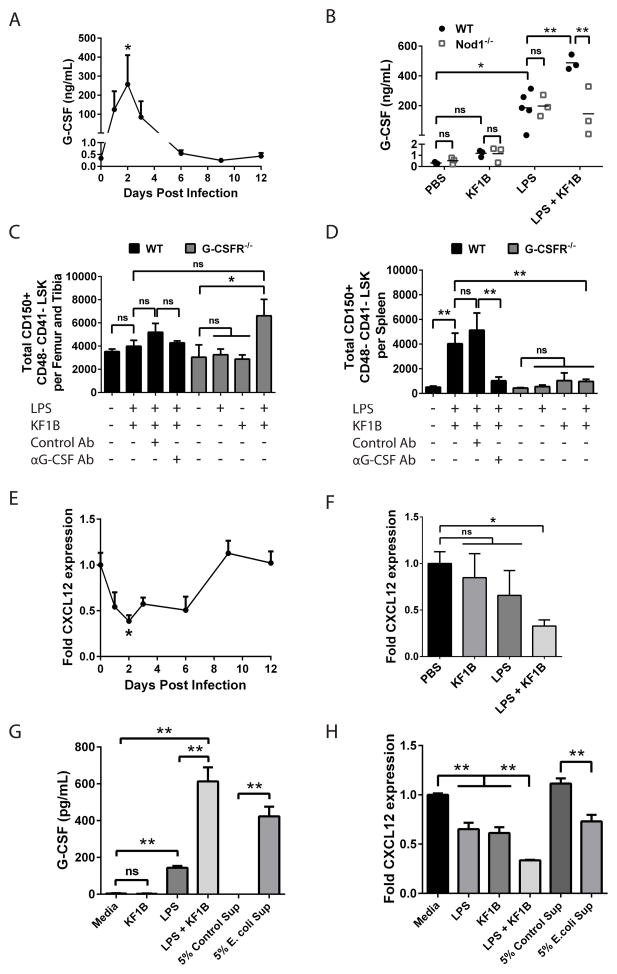
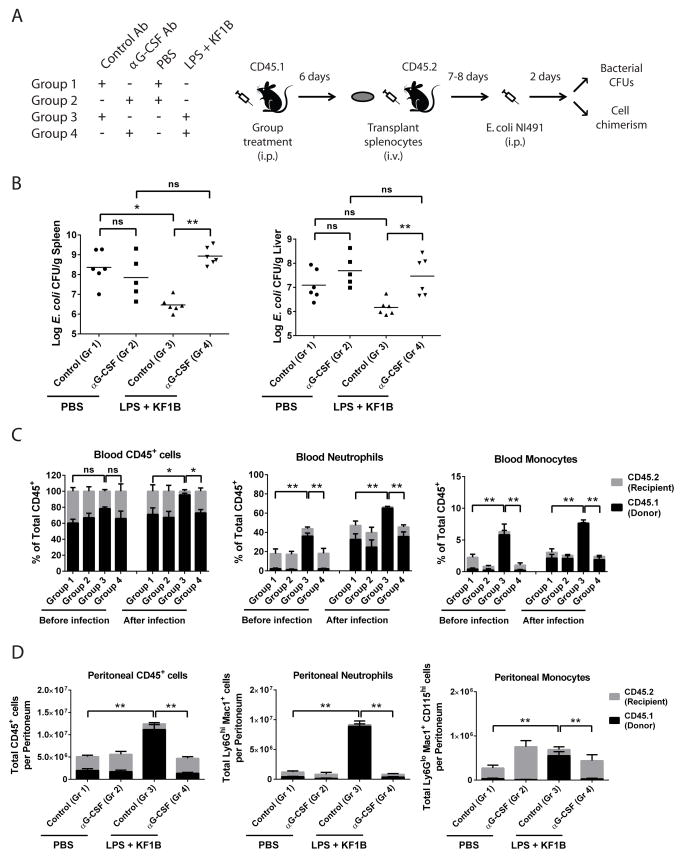
Adult hematopoietic stem cells (HSCs) are maintained in specialized niches within the bone marrow under steady-state conditions and mobilize for extramedullary hematopoiesis during periods of stress such as bacterial infections. However, the underlying mechanisms are unclear. We show that systemic infection of mice with Escherichia coli, commonly associated with bacteremia in humans, mobilizes functional HSCs to the spleen. Accumulation of splenic HSCs (CD150+CD48-Lin(-/low)Sca1+cKit+) was diminished in TLR4-deficient and RIPK2-deficient mice, implicating TLRs and cytosolic NOD1/NOD2 signaling in the process. Accordingly, dual stimulation of NOD1 and TLR4 in radio-resistant cells alone was sufficient to mobilize HSCs, while TLR4 expression on HSCs was dispensable. Mechanistically, TLR4 and NOD1 synergistically induced granulocyte colony-stimulating factor (G-CSF), which was required for extramedullary HSC accumulation. Mobilized HSCs and progenitor cells gave rise to neutrophils and monocytes and contributed to limiting secondary infection.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Nod-like receptors have a grip on stem cells.Cell Host Microbe. 2014 Jun 11;15(6):659-61. doi: 10.1016/j.chom.2014.05.017. Cell Host Microbe. 2014. PMID: 24922568
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
